Михаил Еремушкин - Двигательная активность и здоровье. От лечебной гимнастики до паркура

- Название:Двигательная активность и здоровье. От лечебной гимнастики до паркура
- Автор:
- Жанр:
- Издательство:Литагент Спорт
- Год:2016
- Город:Москва
- ISBN:978-5-9907239-7-9
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Михаил Еремушкин - Двигательная активность и здоровье. От лечебной гимнастики до паркура краткое содержание
Издание рассчитано на специалистов в области физической культуры и самый широкий круг читателей, интересующихся вопросами здорового образа жизни.
Двигательная активность и здоровье. От лечебной гимнастики до паркура - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Миотатический рефлекс (от греч. myos – мышца, tatis – напряжение) вызывается растяжением мышцы, что приводит к увеличению длины мышечного веретена и, вследствие этого, к повышению частоты генерирования потенциала действия в сенсорном (афферентном) нервном волокне. Синонимы: рефлексы Лидделла – Шеррингтона, Ферстера – Альтенбургера (сокращение мышцы-антагониста паретичной мышцы при пассивном растяжении или активном напряжении последней).
В 1905 г. немецкий биолог Якоб фон Икскюль (1864–1944) впервые продемонстрировал существование цепных миотатических рефлексов на офиурах (змеехвостках) – разновидности морских звезд. Их тело имеет звездообразную форму с очень подвижными членистыми лучами, которые представляют собой ряд позвонков, соединенных мышцами. Икскюль закреплял изолированный луч офиуры горизонтально в штативе так, чтобы его конец свободно свисал, изогнувшись вниз, и наносил на него раздражение. При этом луч всегда отклонялся в сторону растянутых мышц, т. е. кверху.
Подобный механизм координации описан также и у низших червей – немартин. Сокращение предыдущего участка ведет к растяжению ближайшего последующего сегмента тела. В растянутом сегменте раздражаются кожно-мышечные рецепторы, инициируя его сокращение.
Цепные координации поперечно-полосатой мускулатуры обнаружены и у млекопитающих. Их существование было наглядно продемонстрировано Р. Магнусом (1962) на хвосте децеребрированной, а затем спинализированной кошки. Животное укладывали на стол спиной кверху, чтобы хвост свободно свисал через его край. При раздражении кончика хвоста он также всегда отклонялся вверх – в сторону растянутых мышц. После того как переднюю часть туловища кошки перемещали в боковое положение, реакция хвоста менялась – при раздражении он двигался в сторону направления передних лап животного.
По данным И.С. Беритова (1966), цепной миотатический рефлекс может развиться как при быстром, так и при медленном растяжении мышцы и сохраняется все время, пока действует растягивающая сила. Он может появляться при растяжении мышцы всего на 0,8 % ее длины покоя.
Нервные механизмы ствола мозга человека существенно обогащают «двигательный репертуар», обеспечивая координацию правильной установки тела в пространстве за счет шейных и лабиринтных рефлексов (Магнус Р., 1962) и нормального распределения мышечного тонуса. Данные рефлекторные связи, по сути, и являются постуральными кинематическими мышечными цепями.
Рефлексы Магнус – Клейна – тонические рефлексы, согласующие постановку туловища и конечностей с положением головы. Эти рефлексы делятся на две большие группы соответственно тому: идет ли дело о сохранении тела и его частей в равновесии или же о реакции на движения. Те рефлексы, которые обусловливают равновесие тела и поддерживают его при стоянии, сидении, лежании, обозначаются как статические рефлексы; те же рефлексы, которыми организм реагирует на активные и пассивные движения и которые частью компенсируют происходящие при этом смещения, называются статокинетическими.
Р. Магнус (1962) выделял непосредственно следующие варианты рефлексов:
1. Местные статические реакции – миотатический ответ на силы притяжения.
2. Сегментарные статические реакции:
а) перекрестный разгибательный рефлекс – возрастание реакции опоры с одной стороны, когда гомолатеральная конечность вынуждена согнуться в ответ на нервный стимул;
б) растяжение конечности на повышение тонуса противоположной конечности – стоя на ногах, сгибая правую конечность, и толкая направо, согнутая конечность разгибается.
Общие статические реакции:
а) тонические рефлексы затылка:
– вращательные – одностороннему вращению головы сопутствует возрастание миотатических рефлексов растяжения конечностей с той стороны, в которую была повернута голова, и расслабления мускулатуры противолежащей конечности;
– разгибательные – разгибание головы ведет к разгибанию передних и расслаблению задних конечностей;
– сгибательные – сгибание головы вперед ведет к сгибанию передних конечностей и разгибанию задних.
б) тонические лабиринтные рефлексы.
Немаловажная роль в координации движений принадлежит мозжечку. Такие качества движения, как плавность, точность, необходимая сила, реализуются с участием мозжечка путем регуляции временны́х, скоростных и пространственных характеристик движения. Животные с удаленными полушариями, но с сохраненным стволом мозга по координации движений, почти не отличаются от интактных.
Однако в управление движениями человека включены также высшие формы деятельности мозга, связанные с сознанием. Полушария мозга (кора и базальные ядра) обеспечивают наиболее тонкие координации движений: двигательные реакции, приобретенные в индивидуальной жизни. Структурными единицами этих двигательных реакций являются динамические мышечные цепи.
Попытки классифицировать варианты динамических мышечных цепей осуществлялись давно. В разное время к ним относили: сухожильно-мышечные меридианы традиционной китайской чжень-цзю-терапии (Сим М., 1993; Петров К.Б., Митичкина Т.В., 2010), миофасциальные меридианы Рольфинга (Рольф И., 1979; Кузнецов О.В., 2009; Майерс Т., 2010), спиральные миоадаптивные цепи (Кадырова Л.А., Сак Н.Н. и др., 1991), мышечные цепочки Вацлава Войта (Войт В., 1958) и др.
Как правило, все авторы предлагаемых вариантов динамических мышечных цепей рекомендовали для достижения гармонии в теле создать сбалансированный тонус по этим цепям, чтобы мышцы находились в состоянии некоего равновесия. Но если по отношению к постуральным кинематическим цепям может идти речь о гармонизации мышечного тонуса, широкая вариативность двигательной активности человека не позволяет говорить о каких-либо закрепленных, строго регламентированных динамических мышечных цепях. Достаточно понаблюдать за атлетами, выполняющими гимнастические упражнения на спортивных соревнованиях, или артистами балета при выполнении сложных сольных партий на сцене (рис. 4).
С 1920–1930 гг. российскими и немецкими учеными, на базе частного раздела «анатомии человека» – «функциональной анатомии», активно стала развиваться наука – «биомеханика движений», которая продемонстрировала, что в разных положениях, при различных двигательных актах под контролем сознания могут включаться абсолютно разные мышечные цепи.
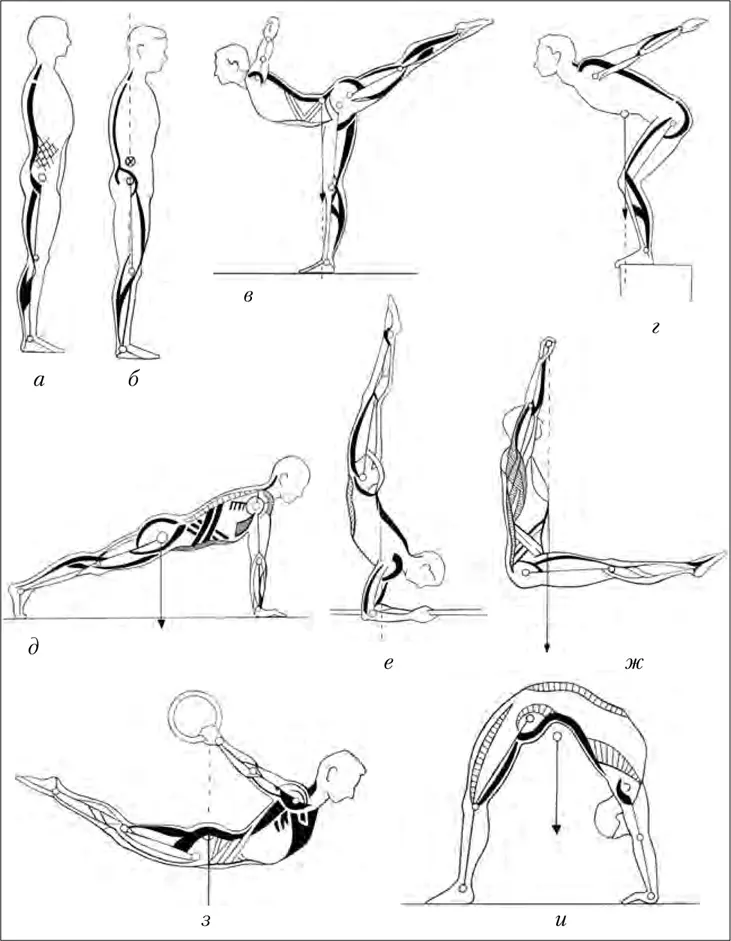
Рис. 4. Мышечные цепи при выполнении спортивных упражнений (по Донскому Д.Д., 1971): а – стойка «Смирно!»; б – «Удобная» стойка; в – равновесие в стойке с опорой на правую ногу; г – старт перед прыжком в воду; д – упор лежа; е – стойка на предплечьях; ж – угол в висе на перекладине; з – горизонтальный вис сзади на кольцах; и – мост на полу. Мышцы, совершающие преимущественно удерживающую работу, обозначены черным цветом, укрепляющую – продольной штриховкой, фиксирующую – поперечной штриховкой
Читать дальшеИнтервал:
Закладка:









