БСЭ БСЭ - Большая Советская Энциклопедия (ДЫ)

- Название:Большая Советская Энциклопедия (ДЫ)
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
БСЭ БСЭ - Большая Советская Энциклопедия (ДЫ) краткое содержание
Большая Советская Энциклопедия (ДЫ) - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Лит.: Дудко П. Н., Сортовое богатство дынь Узбекистана, Таш., 1956; Пангало К. Т., Дыни, Киш., 1958; Юрина О. В., Селекция и семеноводство тыквенных культур, М., 1966.
В. Ф. Белик.
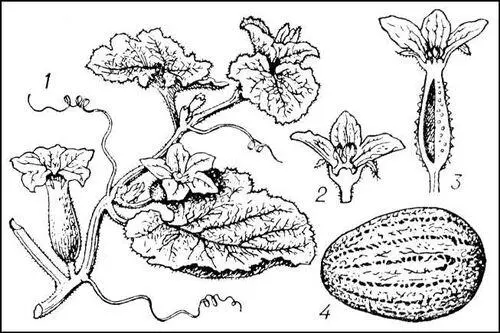
Дыня: 1 — общий вид растения; 2 — тычиночный цветок; 3 — пестичный цветок; 4 — плод.
Дыравка
Дыра'вка,род травянистых растений семейства мальвовых, известных в садоводстве под названием малопе .
Дырка
Ды'рка(в зонной теории твёрдого тела ), не занятое электроном энергетическое состояние, например в валентной зоне полупроводника . Д. ведут себя как частицы ( квазичастицы ) с положительным зарядом, равным по абсолютной величине заряду электрона, и являются наряду с электронами носителями тока в полупроводниках (дырочная электропроводность).
Лит . см. при ст. Твёрдое тело , Полупроводники .
Дыхание
Дыха'ние,совокупность процессов, которые обеспечивают поступление в организм кислорода и выделение из него углекислого газа (внешнее Д.) и использование кислорода клетками и тканями для окисления органических веществ с освобождением содержащейся в них энергии, необходимой для жизнедеятельности ( тканевое дыхание , клеточное Д.). Бескислородный путь освобождения энергии свойствен только небольшой группе организмов — так называемым анаэробам (см. Брожение ); в ходе эволюции освобождение энергии в результате Д. стало у подавляющего большинства организмов главным процессом, а анаэробные реакции сохранились в основном как промежуточные этапы обмена веществ .
Д. животных и человека.У простейших, губок, кишечнополостных и некоторых др. организмов О 2диффундирует непосредственно через поверхность тела. С усложнением организации и увеличением размеров тела появляются специальные дыхания органы , а также система кровообращения, в которой циркулирует жидкость — кровь или гемолимфа, содержащая вещества, способные связывать и переносить O 2и CO 2(см. Гемоглобин ). У насекомых O 2поступает в ткани из системы воздухоносных трубочек — трахей . У водных животных, использующих растворённый в воде O 2, органами Д. служат жабры , снабжённые богатой сетью кровеносных сосудов. В этом случае O 2, растворённый в воде, диффундирует в кровь, циркулирующую в сосудах жаберных щелей. У многих рыб значительную роль играет кишечное Д., при котором воздух заглатывается и O 2поступает в кровь через кровеносные сосуды кишечника; некоторую роль в Д. рыб играет также плавательный пузырь; у многих обитающих в воде животных обмен газов (главным образом СО 2) происходит и через кожу. У сухопутных животных внешнее Д. обеспечивается преимущественно лёгкими . У земноводных и многих др. животных наряду с этим функционирует кожное Д. У птиц существенное значение имеют сообщающиеся с лёгкими воздушные мешки , которые изменяются в объёме при летательных движениях и облегчают Д. в полёте. У земноводных и пресмыкающихся воздух в лёгкие нагнетается движениями мышц дна рта. У птиц, млекопитающих и человека внешнее Д. обеспечивается ритмической работой дыхательных мышц (главным образом диафрагмы и межрёберных мышц), координируемой нервной системой. При сокращении этих мышц объём грудной клетки увеличивается и происходит растяжение находящихся в ней лёгких; поэтому возникает разность между атмосферным и внутрилёгочным давлением и воздух поступает в лёгкие (вдох). Выдох может быть пассивным — за счёт спадения растянутой во время вдоха грудной клетки, а вслед за ней и лёгких; активный выдох обусловлен сокращением некоторых групп мышц. Количество воздуха, поступающее в легкие за 1 вдох, называется дыхательным объёмом (см. Лёгочные объёмы ).
При Д. дыхательная мускулатура преодолевает эластичное сопротивление, связанное с упругостью грудной клетки, тягой лёгких и поверхностным натяжением альвеол. Последнее, однако, значительно снижается под влиянием поверхностно активного вещества, вырабатываемого клетками альвеолярного эпителия; поэтому альвеолы при выдохе не спадаются, а при вдохе легко расширяются. Чем выше эластичное сопротивление, тем труднее растягиваются грудная клетка и лёгкие; при глубоком Д. работа дыхательной мускулатуры, затрачиваемая на его преодоление, резко возрастает. Неэластичное сопротивление Д. обусловлено главным образом трением воздуха при его движении по носовым ходам, гортани, трахее и бронхам. Оно зависит от скорости потока воздуха во время Д. и от его характера. При спокойном Д. поток близок к ламинарному (линейному) в прямых участках воздухоносных путей и к турбулентному (вихревому) в местах разветвления или сужения. С увеличением скорости потока (при форсированном Д.) турбулентность возрастает и для продвижения воздуха требуется более высокая разность давлений, а следовательно, и увеличение работы дыхательных мышц. Неравномерное распределение сопротивления движению воздуха по дыхательным путям приводит к тому, что поступление воздуха в разные группы лёгочных альвеол происходит неравномерно; такая разница в вентиляции особенно значительна при лёгочных заболеваниях.
Количество воздуха, вентилирующее лёгкие в 1 мин , называется минутным объёмом дыхания (МОД). МОД равен произведению дыхательного объёма на частоту Д. (число дыхательных движений в 1 мин , равное у человека примерно 15—18) и составляет у взрослого человека в покое 5—8 л/мин . Только часть МОД (около 70%) участвует в обмене газов между вдыхаемым и альвеолярным воздухом, эту часть называют объёмом альвеолярной вентиляции; остальная часть МОД используется для «промывания» так называемого мёртвого, или вредного, пространства дыхательных путей, в котором к началу выдоха сохраняется наружный воздух, заполнивший его в конце предшествовавшего вдоха (объём мёртвого пространства около 160 мл ). Вентиляция альвеол обеспечивает постоянство состава альвеолярного воздуха . Парциальное давление O 2( p O2) и CO 2( p CO2) в альвеолярном воздухе колеблется в очень узких пределах и составляет для О 2около 13 кн/м 2 (100 мм рт. cт. ) и для СО 2около 5,4 кн/м 2 (40 мм рт. ст. ).
Обмен газов между альвеолярным воздухом и венозной кровью, поступающей в капилляры лёгких, осуществляется через альвеоло-капиллярную мембрану, общая поверхность которой очень велика (у человека около 90 м 2 ). Диффузия O 2в кровь обеспечивается разностью парциальных давлений O 2в альвеолярном воздухе и в венозной крови (8—9 кн/м 2 , или 60—70 мм рт. ст. ). CO 2, приносимый кровью из тканей в связанной форме (бикарбонаты, соли угольной кислоты и карбгемоглобин), освобождается в капиллярах лёгких при участии фермента карбоангидразы и диффундирует из крови в альвеолы; разность p CO2между венозной кровью и альвеолярным воздухом составляет около 7 мм рт. ст. Способность альвеолярной стенки пропускать O 2и CO 2, так называемая диффузионная способность лёгких, очень велика: в покое она составляет в 1 мин примерно 30 мл O 2на 1 мм разности p O2между альвеолярным воздухом и кровью; для CO 2эта величина во много раз больше. Поэтому парциальное давление газов в оттекающей из лёгких артериальной крови успевает приблизиться к их давлению в альвеолярном воздухе. Переход O 2в ткани и удаление из них CO 2также происходят путём диффузии, т.к. p O2в тканевой жидкости 2,7—5,4 кн/м 2 (20—40 мм рт. ст. ), а в клетках ещё ниже, а p CO2в клетках может достигать 60 мм рт. cт. (см. рис. ).
Читать дальшеИнтервал:
Закладка: