Владислав Солоухин - Размышления и споры о вирусах

- Название:Размышления и споры о вирусах
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:1989
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Владислав Солоухин - Размышления и споры о вирусах краткое содержание
Размышления и споры о вирусах - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
За неспецифической стадией адсорбции наступает высокоспецифическая, когда белки поверхности вируса (так называемые прикрепительные) соединяются со специфическими группировками (рецепторами) на плазматической мембране клетки, если, конечно, такие рецепторы на этих клетках имеются. Для вируса гриппа, например, на слизистой верхних дыхательных путей они есть, а на коже нет. Именно поэтому грипп — респираторная инфекция, а полиомиелит — кишечная (во всяком случае — по методу заражения или, как говорят эпидемиологи, по входным воротам инфекции).
Мы не будем рассматривать структуру рецепторов на разных клетках и свойства прикрепительных белков представителей разных рибо- и дезоксирибовирусов. Скажем лишь: что в своей совокупности эти сведения достаточно точно и полно объясняют явление тропизма вирусов к одним тканям, устойчивость тех или иных клеток и вирусам и, наконец, видовую устойчивость или, наоборот, чувствительность к вирусным инфекциям. Почему, например, пикорнавирусы (возбудители полиомиелита и полиомиелитоподобных инфекций) поражают только приматов, а тогавирусы (возбудители клещевого и комариного энцефалитов) — и млекопитающих, и членистоногих, и человека? Да потому, что соотношение структуры прикрепительных вирусных белков и клеточных рецепторов не позволяет пикорнавирусам выйти за пределы отряда приматов, а тогавирусам дает возможность легко перешагнуть границы типов и проникать в клетки как позвоночных, так и беспозвоночных.
Итак, адсорбция прошла. Что же дальше? Дальше — проникновение вируса в клетку. Исторически сложилось представление о том, что вирусы могут проникать в клетки либо путем так называемого виропексиса, либо путем слияния вирусной и клеточной мембран. Вряд ли правильно говорить либо — либо. По-видимому, оба эти механизма не столько исключают, сколько дополняют друг друга. Понятно также, что каждая система вирус-клетка характеризуется своим индивидуальным способом проникновения нарушителя границы клеточного государства.
Современные руководства по вирусологии буквально переполнены описанием подробностей каждого такого отдельного разбоя, и только в некоторых из них содержатся отдельные попытки прописать лекарства, защищающие клетки от гибели. Мы имеем в виду химиотерапевтические средства, к сожалению, крайне немногочисленные в вирусологии. Но об этом — в другом месте. Сейчас в нескольких словах охарактеризуем самое главное — проникновение вируса в клетку.
Термин "виропексис" предложил в 1948 году видный вирусолог австралиец венгерского происхождения Ф. де Сент-Гроот. Это не что иное, как разновидность пиноцитоза (или эндоцитоза), то есть обычного механизма поступления в клетку из внешней среды питательных и регуляторных белков, гормонов и других высокомолекулярных веществ. В отдельных участках цитоплазмы, в особых ямках, где располагаются специфические рецепторы, после взаимодействия с вирусом возникают выпячивания (инвагинации). Затем эти выпячивания замыкаются и образуются внутриклеточные вакуоли, содержащие в себе проникшее в клетку вещество, в частности, вирус, если это был он.
Вакуоли сливаются между собой, образуя крупные внутриклеточные полости, соединяющиеся затем с лизосомами. Вы помните об эти стражах постоянства внутренней среды клеток, наполненных разнообразными ферментами? Теперь мы их видим в работе: содержимое лизосом изливается в вакуоли, и начинается эффективная ферментативная переработка. Белки разлагаются до аминокислот, нуклеиновые кислоты до отдельных мононуклеотидов и т. д. Казалось бы, лизосомы должны в порошок растереть непрошеных гостей, ведь вирусы — типичные нуклеопротеиды! Но этого не происходит. Напротив, лизосомы своими ферментами помогают вирусной мембране слиться с клеточной, выстилающей внутреннюю полость вакуоли. Вот почему мы говорили о том, что механизмы виропексиса и слияния мембран вируса и клетки не противоречат, а дополняют друг друга.
Итак, мембраны слиты! В одних системах вирус — клетка это известно до мельчайших подробностей, в других — только начинает изучаться. Например: какой из вирусных белков является белком слияния? По-настоящему он идентифицирован только у пара- и ортомиксо-вирусов. Так, у вируса кори или парагриппа (оба — парамиксовирусы) им является один из двух гликопротеидов, находящихся на поверхности вирусной частицы. У вируса гриппа (ортомиксовирус) функцию белка слияния выполняет малая субъединица гемагглютинина — важнейшего поверхностного антигена этого возбудителя. Так или иначе, но этот процесс происходит при проникновении в клетки любых вирусов.
Итак, все сказанное подводит нас к главнейшему событию, без которого не может начаться вирусная инфекция — "раздеванию", высвобождению вирусного генома от белкового покрытия. Условия для такого раздевания создаются всеми предшествующими, описанными нами процессами: связыванием со специфическими рецепторами, проникновением в клетку, слиянием мембран.
Не отвлекаясь на частности (а великое множество их уже известно и еще более неизвестно), можно сказать, что вирус, проходя внутрь клетки, раздевается постепенно, сбрасывая по мере своего взаимодействия с клеткой одну одежду за другой, стремясь обнажиться до такой степени, которая позволит ему начать смертельный для клетки "танец": для одних вирусов такими продуктами являются сердцевины вирионов, в других — нуклеокапсиды, в третьих — нуклеиновые кислоты. Но во всех случаях критерием достаточной для начала инфекции степени раздевания вируса является способность начать транскрипцию.
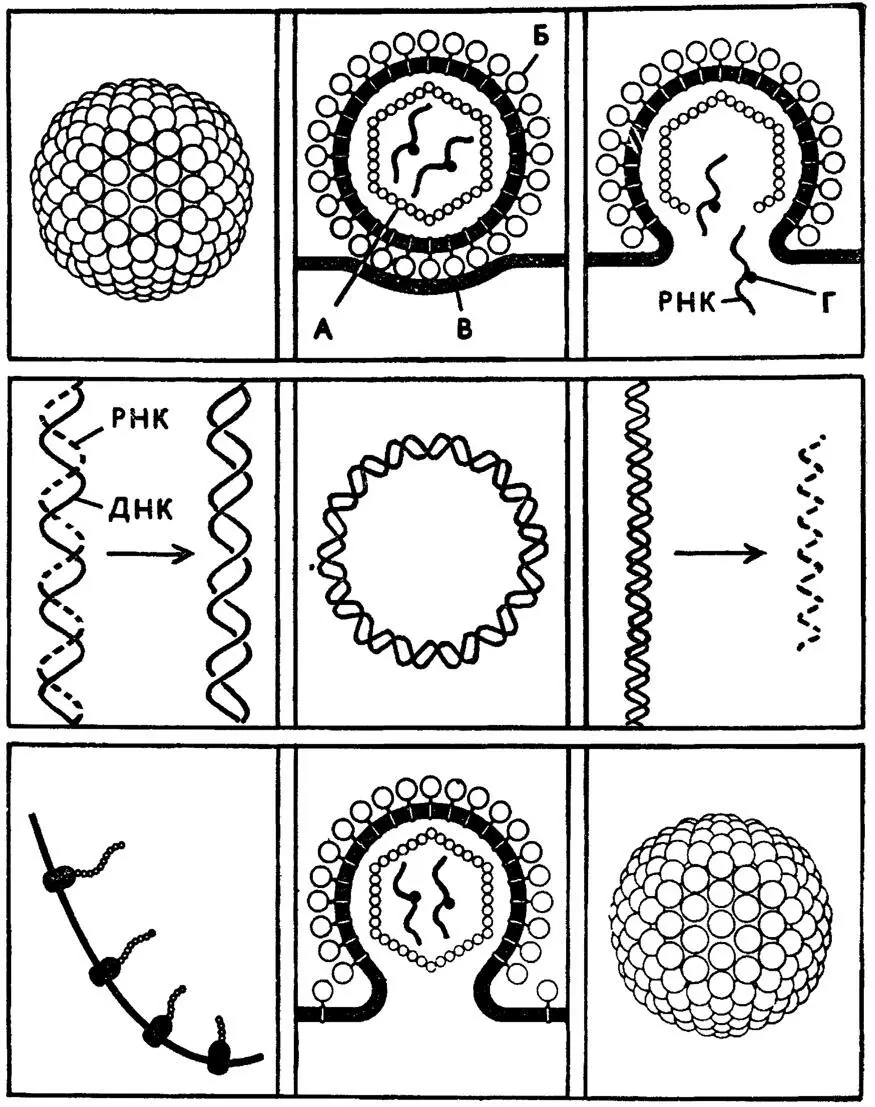
Рис. 5. Жизненный цикл ретровируса. А — белок сердцевины; Б — гликопротеин оболочки; В — клеточная мембрана; Г — обратная транскрипция
Нам не нужно здесь объяснять, что такое транскрипция — мы говорили об этом выше, путешествуя по клетке. Напомним, что центральная догма молекулярной биологии клетки выражается формулой: ДНК (транскрипция)→РНК (трансляция)→белок.
То, как реализуют свою генетическую информацию вирусы, называют — стратегией вирусного генома (!). (Заметим: это не наше выражение, мы заимствуем его из обширной литературы по молекулярной вирусологии.) Стратегия генома ДНК-содержащих вирусов в отношении синтеза иРНК та же самая, что и у клетки (ДНК→РНК→белок). Стратегия та же! А тактика?
Тактика различна у разных групп вирусных агентов. Большинство ДНК-содержащих вирусов, во всяком случае все, репродукция которых происходит в ядре (паповавирусы, аденовирусы, вирусы герпеса), используют для своей транскрипции клеточный фермент полимеразу (точнее: ДНК-зависимую РНК-полимеразу). Именно она на матрице вирусной ДНК строит комплементарную ей информационную иРНК. Вот она — молекулярная структура вирусного паразитизма, молекулярное выражение разбойного поведения вируса, проникшего в клетку-государство и использующего его "правительственный аппарат" для своих злодейских целей!
Читать дальшеИнтервал:
Закладка:





