Изот Литинецкий - Беседы о бионике

- Название:Беседы о бионике
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:1968
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Изот Литинецкий - Беседы о бионике краткое содержание
Беседы о бионике - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Тело нервной клетки в поперечнике обычно меньше 0,1 мм. Объем крупного нейрона составляет примерно 0,001 мм 3. Дендриты имеют диаметр порядка 0,01 мм и длину от долей миллиметра до десятков сантиметров. Длина аксона нервных клеток человека колеблется от долей миллиметра до 1,5 м (при толщине около 0,025 мм).
По современным представлениям, в основе функции реального нейрона лежат электрохимические процессы. Его мембрана состоит из четырех мономолекулярных слоев (белок — липоид — липоид — белок) общей толщиной около 10 -6см. Нейрон в состоянии покоя имеет следующие электрические параметры: разность потенциалов 70 мв, удельное сопротивление 0,4 · 10 12ом · см и емкость 1 мкф/см 2(данные измерений между наружными поверхностями мембраны).
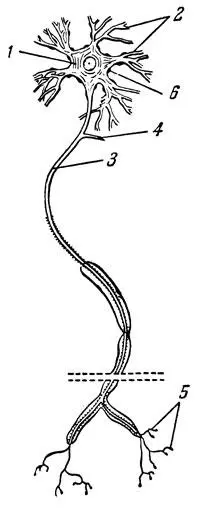
Рис. 1. Схематическое изображение нейрона (нервной клетки)
Нервы в организме играют роль линий связи между рецепторными нервными клетками с чувствительными окончаниями, воспринимающими информацию, скоплениями нейронов, предназначенных для обработки информации, и исполнительными, или эффекторными, клетками, обеспечивающими соответствующие реакции отдельных органов или участков организма. Все эти элементы, воспринимающие, передающие, перерабатывающие и выдающие управляющую информацию, образуют нервную систему. Функция нервной системы основана на процессах возбуждения и торможения. Возбуждение возникает под влиянием электрических, тепловых, химических и механических раздражений и распространяется по нервным волокнам в виде электрических импульсов. Импульс, возникающий в нейроне, распространяется по аксону без затухания и с постоянной скоростью, примерно равной a VD, где а — постоянная величина, a √D — диаметр аксона. Таким образом, чем толще аксон, тем больше скорость распространения импульсов. Эта скорость неодинакова у различных организмов. У человека она не превышает 120 м/сек, у собаки она составляет 83,3 м/сек, у улитки — 0,05 — 0,4 м/сек.
Посмотрим теперь, как "работает" нейрон. Он может находиться в двух состояниях: возбуждения и торможения. Различные части нейрона несут разные функции. Дендриты служат входами, по которым к телу клетки подводятся импульсы раздражения, а аксоны — выходами, по которым передается возбуждение на другие клетки. В передаче нервными волокнами возбуждения велика роль синапсов, т. е. мест перехода возбуждения от одной нервной клетки к другой. Синапсы обладают односторонней проводимостью, т. е. возбуждение передается только с окончаний аксона одного нейрона на дендриты и тело клетки другого нейрона (на крупных нейронах насчитывается до 1000 синапсов). Кроме односторонней проводимости синапс характеризуется еще одним интересным свойством: в нем происходит замедление проведения возбуждения, т. е. замедляется передача раздражения. Это называется синаптической задержкой. Прохождение возбуждения через синапс как бы подготавливает почву и облегчает прохождение через него следующего возбуждения.
Нейрон имеет множество входов, а выход у него только один. Входной импульс может быть возбуждающим или тормозящим; он может иметь самые различные параметры; выходной же сигнал каждого нейрона представляет собой импульс, амплитуда и длительность которого постоянны. Примечательно, что нейрон срабатывает только при определенном уровне входного воздействия, называемом пороговым. Если раздражение, поступающее на один из входов, ниже, этого уровня, то нейрон продолжает оставаться невозбужденным. Если оно выше этого критического уровня, то нейрон переходит в возбужденное состояние, при котором энергия из тела клетки передается в виде стандартного импульса в аксон.
В течение некоторого времени после разряда нейрона входные сигналы не вызывают его возбуждения.
Это объясняется резким повышением порогового уровня при генерировании нейроном выходного сигнала. В дальнейшем за время, называемое временем восстановления, пороговый уровень понижается до прежней величины. Состояние нейрона после его срабатывания характеризуется возбудимостью — величиной, обратной пороговому уровню. Кривая зависимости возбудимости нейрона от времени приведена на рис. 2, где по оси абсцисс отложено время, прошедшее после срабатывания нервной клетки, а по оси ординат — величины, обратные пороговому уровню (в процентах от нормального значения).
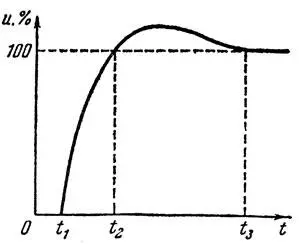
Рис. 2. Кривая зависимости возбудимости нейрона от времени (по Л. И. Крайзмеру)
Как видно из графика, следующий разряд нейрона может произойти не ранее, чем через некоторый интервал времени от 0 до t 1. Такой интервал, называемый периодом абсолютной рефрактерности, длится от 0,4 до 2 мсек. В течение этого времени пороговой уровень нейрона как бы оказывается равным бесконечности, а возбудимость его равна нулю. Считают, что время пониженной возбудимости нейрона от момента его срабатывания до восстановления нормального порога определяется необходимостью восстановить энергию клетки, затраченную на подготовку и генерирование выходного импульса. После окончания периода полной невосприимчивости происходит постепенное (в течение нескольких десятков миллисекунд) снижение порогового уровня до нормальной величины. Этот период (от момента t 1до t 2) носит название периода относительной рефрактерности. За ним следует фаза повышенной возбудимости нейрона (от момента t 1до t 3), во время которой он может сработать и при воздействии возбуждения ниже порогового уровня. Наконец, в некоторый момент времени t 3восстанавливается нормальная возбудимость нейрона.
Но всем этим динамика нейрона не исчерпывается. Раздражения, уровни которых недостаточны для возбуждения, но действующие на несколько входов одновременно, суммируются и возбуждают нейрон — это так называемое пространственное суммирование. Раздражения с уровнями ниже порога, но следующие одно за другим через короткие промежутки времени, тоже суммируются и возбуждают нейрон. Здесь происходит последовательное суммирование, или суммирование во времени. В механизме возбуждения нейрона часто имеют место одновременно оба процесса — суммирование раздражений как в пространстве, так и во времени. При этом роль предыдущих импульсов возбуждения становится тем меньше, чем больше времени прошло после их появления. Происходит как бы затухание их следов, которое подчиняется экспоненциальному закону.
Как отмечалось выше, импульсы раздражения могут быть возбуждающими или тормозящими и они могут поступать на разные входы нейрона одновременно. Если их алгебраическая сумма превышает пороговый уровень, то нейрон возбуждается и выдает импульс.
Читать дальшеИнтервал:
Закладка:








