Кирилл Еськов - Доказательства эволюции

- Название:Доказательства эволюции
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:2010
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Кирилл Еськов - Доказательства эволюции краткое содержание
). Отличия файла от оригинала минимальны. В файле отсутствуют по понятным причинам несколько видеороликов, которые имеются в оригинале, но все изображения и гиперссылки сохранены. Кроме того, в файле отсутствует самая последняя глава — фрагменты из книги Ф. Коллинза.
Доказательства эволюции - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
«ЭКОЛОГИЧЕСКАЯ СОРТИРОВКА». Еще один хитроумный зигзаг антиэволюционной мысли породил «теорию», согласно которой «сортировка» ископаемых во время Потопа объясняется тем, что разные животные с разной эффективностью «спасались» от бедствия: низшие формы погибли первыми, тогда как высшие продержались дольше — например, потому, что были способны убежать в горы. Что касается растений, не способных куда-либо убежать, то их закономерное распределение в земных слоях противники эволюции объясняют, видимо, тем, что низшие растения росли ниже, а высшие — выше над уровнем моря (картина, имеющая мало общего с реальностью). Эта «теория» еще смехотворнее и беспомощнее, чем «модель гидродинамической сортировки». Она рассчитана исключительно на людей, абсолютно не знакомых с реальными данными стратиграфии и палеонтологии. Например, как объяснить с точки зрения этой теории, что и пыльца и споры наземных растений в осадочных породах тоже распределяются в соответствии с общим правилом? В докембрийских и кембрийских отложениях вообще нет пыльцы и спор наземных растений. Первые споры наземных мохообразных появляются в отложениях середины ордовикского периода (около 465 млн лет назад). Споры первых сосудистых растений известны с конца ордовика (450–444 млн лет назад) (см.: Палеонтологи уточнили время и место появления сосудистых растений). И так далее, вплоть до появления пыльцы цветковых растений (и самих окаменевших цветковых растений) в меловом периоде (немногим более 100 млн лет назад). Прекрасно известно, что пыльца и споры наземных растений далеко разносятся ветром, тонут в морях и поэтому повсеместно присутствуют в морских отложениях, образующихся в наши дни. Так же обстояло дело и в прошлом. Поэтому «споро-пыльцевой» анализ — один из надежнейших методов биостратиграфии, позволяющий с большой точностью определять возраст любых осадочных отложений, как морских, так и континентальных. «Теория экологической сортировки» не в состоянии объяснить стратиграфическое распространение пыльцы и спор. То же самое со всей очевидностью относится к таким категориям ископаемых, как: 1) окаменевшие следы (ихнофоссилии), оставленные древними организмами на морском дне и на мягкой почве континентов, 2) морской планктон (радиолярии, фораминиферы, кокколитофориды и т. д.) — какая у них-то могла быть экологическая сортировка? 3) древние озерные отложения; хороший пример — отложения юрского озера Каратау, где есть разнообразные остатки юрских позвоночных (в т. ч. мелких птерозавров), насекомых, растений, но нет ни морских организмов, ни окаменелостей, принадлежащих животным и растениям других эпох. Причем тут всемирный Потоп? Объяснить всю совокупность палеонтолого-биостратиграфических данных всемирным Потопом абсолютно немыслимо, если иметь об этих данных сколько-нибудь полное представление.
КАТАСТРОФИЗМ. На ранних этапах развития исторической геологии и палеонтологии, когда было выделено еще очень небольшое число геохронологических интервалов, наблюдаемую картину пытались объяснить при помощи теории катастроф(Бог несколько раз уничтожал свое Творение глобальными катастрофами, а потом творил заново). Но сегодня глобальные (узнаваемые по всему миру) геохронологические интервалы, каждый со своим уникальным комплексом окаменелостей, уже исчисляются сотнями, а преобразования животного и растительного мира в последовательностях слоев явно выглядят во многих случаях вполне постепенными. Даже самые ярые антиэволюционисты не могут вообразить, зачем могло понадобиться Богу сотни раз уничтожать жизнь, а потом каждый раз воссоздавать ее лишь в слегка измененном виде. Развитие исторической геологии и палеонтологии, таким образом, опровергло катастрофизм. Тем не менее в истории Земли действительно были периоды массовых вымираний, сыгравшие значительную роль в эволюции биосферы (см.: Экологические кризисы, их причины и эволюционная роль).
Биостратиграфия
Как и во многих других случаях, для того, чтобы получить верное представление о степени убедительности палеонтологических доказательств эволюции, необходимо прежде всего осознать масштабпроисходящего, всю громадность объема накопленных данных. Для начала рассмотрим современную геохронологическую шкалу — результат многолетних трудов многотысячной международной армии геологов ( См. также: Стратиграфия и геохронология ).
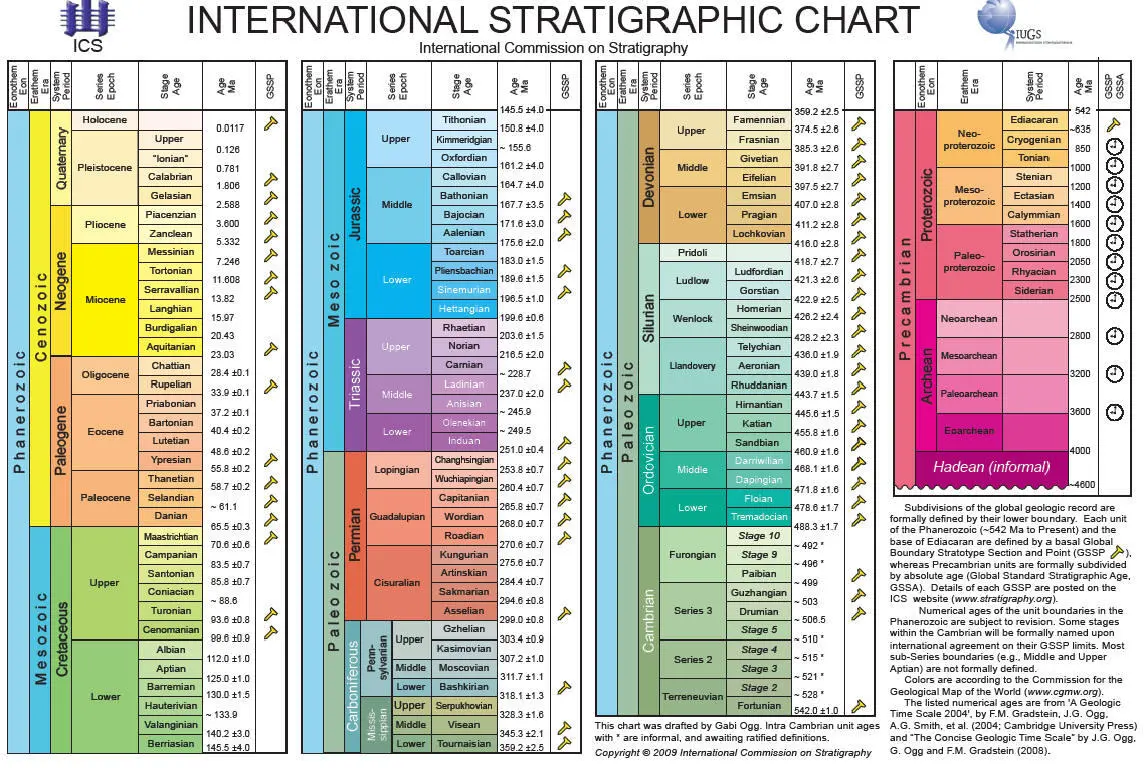
{Ссылка на более крупное ищображение: http://evolbiol.ru/evidence05_files/eviden1.jpg}
На этой шкале выделено 115 временных (геохронологических) интервалов. Это «международные» интервалы, то есть такие, которые удается опознать по всему миру. В каждом регионе существуют еще и местные (региональные) стратиграфические шкалы, которые, как правило, намного более детальны. Однако многие региональные слои не удается точно соотнести («скоррелировать», как говорят геологи) со слоями, известными в других регионах. Поэтому общее количество выделяемых слоев на международной шкале значительно меньше, чем на региональных шкалах. Почему же не все региональные слои удается скоррелировать? Дело в том, что корреляция слоев в большинстве случаев осуществляется по комплексам окаменелостей. А в разных регионах во все времена, как и сегодня, жили разные организмы. Для межрегиональной корреляции удобнее всего использовать такие ископаемые организмы, которые: 1) были распространены всесветно (такие организмы называют «космополитами»), 2) были достаточно многочисленными и неплохо сохраняющимися в ископаемом состоянии (для этого весьма желательно наличие скелета), 3) существовали недолго (иначе они будут встречаться во многих слоях, и их нельзя будет использовать для идентификации конкретного слоя). Ископаемые организмы, удовлетворяющие этим условиям, называют «руководящими ископаемыми», и они играют ключевую роль в стратиграфии. Как правило, роль руководящих форм играют группы организмов, для которых была характерна наиболее быстрая эволюция, быстрая смена видового состава. Как видим, стратиграфия — это наука, в которой геология теснейшим образом переплелась с эволюционным учением. Очень хорошими руководящими ископаемыми являются споры и пыльца растений, а также разнообразные мелкие планктонные организмы (например, фораминиферы, радиолярии, диатомовые водоросли, кокколитофориды), а во многих случаях и более крупные животные (трилобиты, аммониты, конодонты, археоциаты, двустворчатые и брюхоногие моллюски, брахиоподы и мн. др.). Каждый интервал геохронологической шкалы, кроме докембрийских (показаны на рисунке справа), характеризуется уникальным комплексом руководящих ископаемых, а также гораздо более обширным комплексом всех прочих ископаемых, по которым коррелировать слои тоже можно, но не так удобно. В докембрии ископаемых мало, и поэтому там стратиграфия базируется в основном на радиометрическом датировании (определении абсолютного возраста) горных пород. В настоящее время уже установилось практически полное согласие между палеонтологическими (основанными на эволюции организмов) и физическими (радиометрическими) методами геологической корреляции. Таким образом, налицо союз трех наук: геологии, эволюционной биологии и физики. Все вместе они дают стройную и непротиворечивую картину постепенного развития жизни на нашей планете.
Читать дальшеИнтервал:
Закладка:



