Кирилл Еськов - Доказательства эволюции

- Название:Доказательства эволюции
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:2010
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Кирилл Еськов - Доказательства эволюции краткое содержание
). Отличия файла от оригинала минимальны. В файле отсутствуют по понятным причинам несколько видеороликов, которые имеются в оригинале, но все изображения и гиперссылки сохранены. Кроме того, в файле отсутствует самая последняя глава — фрагменты из книги Ф. Коллинза.
Доказательства эволюции - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Как говорилось в разделе «Наблюдаемая эволюция», ученым удается, несмотря на все трудности и низкие вероятности, реально наблюдать процесс видообразования в природе и эксперименте. Но можем ли мы точно так же пронаблюдать появление нового семейства? По-видимому, не можем, и вовсе не потому, что таких событий в природе не бывает, а просто по определению, в силу того обстоятельства, что никакая новая форма, выведенная в лаборатории или возникшая на наших глазах в природе, не будет признанановым семейством. Биологи не станут выделять ее в новое семейство именно потому, что все произошло так быстро. Нравится нам это или нет, но ранг семейства биологи присваивают группе только в ретроспективе, когда группа уже «набрала силу», накопила запас изменчивости, разделилась на роды и виды, одним словом, «доказала» всей своей эволюционной историей, что заслуживает семейственного ранга. Даже если в результате «макромутации» возникнет необычная форма с измененным планом строения, потребуются миллионы лет, чтобы выяснить, можно ли эту форму считать родоначальником нового семейства, или это был просто бесперспективный уродец, обреченный на вымирание, или аберрантный представитель предкового семейства, от которого так и не произошло ничего принципиально нового.
Поэтому мы можем реально наблюдать «настоящие» макроэволюционные события только в ископаемой летописи, когда удается найти хорошие филогенетические ряды, соединяющие предковый таксон (например, лопастеперых рыб) с таксоном-потомком (например, первыми бесспорными тетраподами), о котором нам уже заранее известно, что от него в будущем действительно произошло что-то большое, важное и принципиально новое (например, все наземные позвоночные).
Рассмотрим эту ситуацию подробнее. Лопастеперые рыбы дали начало первым наземным позвоночным в конце девонского периода. Это «макроэволюционное событие» довольно детально документировано в ископаемой летописи. Переход от рыб к тетраподам растянулся примерно на 20–25 млн лет (385–360 млн лет назад) и состоял из множества последовательных мелких шажков, ни один из которых по отдельности никак «не тянет» на статус макроэволюционного.
Легкие (и, соответственно, два круга кровообращения) были, судя по палеонтологическим данным, у большинства или даже у всех девонских лопастеперых рыб. Они сохранились и у некоторых современных рыб. В девоне было понижено содержание кислорода в атмосфере, а доля углекислого газа, наоборот, была повышена по сравнению с современностью. Поэтому в воде, особенно на мелководье, рыбам было трудно дышать одними жабрами. Они заглатывали воздух, как это делают и современные рыбы, и у них образовались впячивания пищевода для удержания пузырей воздуха. Эти впячивания и есть легкие. Они выполняли сразу две функции — примитивных легких и примитивного плавательного пузыря. В дальнейшем у большинства костных рыб эти древние легкие стали отделяться от пищевода и превратились в «настоящий» плавательный пузырь. Генетический анализ показал, что из современных рыб ближайшими родственниками тетрапод являются двоякодышащие. У двоякодышащих рыб есть и легкие, и два круга кровообращения, и трехкамерное сердце — совсем как у амфибий, которые относятся к настоящим тетраподам.
По сути дела, основное, что изменилось при переходе от рыб к тетраподам — это строение конечностей. Но конечности (плавники) девонских лопастеперых рыб не так уж сильно отличались по своему строению от лап примитивных тетрапод. А главное, можно в деталях проследить изменения конечностей в эволюционном ряду, соединяющем рыб и тетрапод. На сегодняшний день этот филогенетический ряд включает около дюжины последовательных переходных форм, из которых можно упомянуть наиболее важные (в порядке убывания рыбьих и накопления тетраподных черт строения): Eusthenopteron, Panderichthys, Tiktaalik,Elpistostege, Livoniana, Elginerpeton, Ventastega, Metaxygnathus, Acanthostega, Ichthyostega, Tulerpeton (жирным шрифтом выделены роды, у которых известно строение конечностей; у остальных конечности сохранились плохо или вовсе пока не найдены). Первый род в этом списке — еще «типичная рыба», последний — бесспорная амфибия. Посередине помещаются все промежуточные этапы этого «макроэволюционного» преобразования.
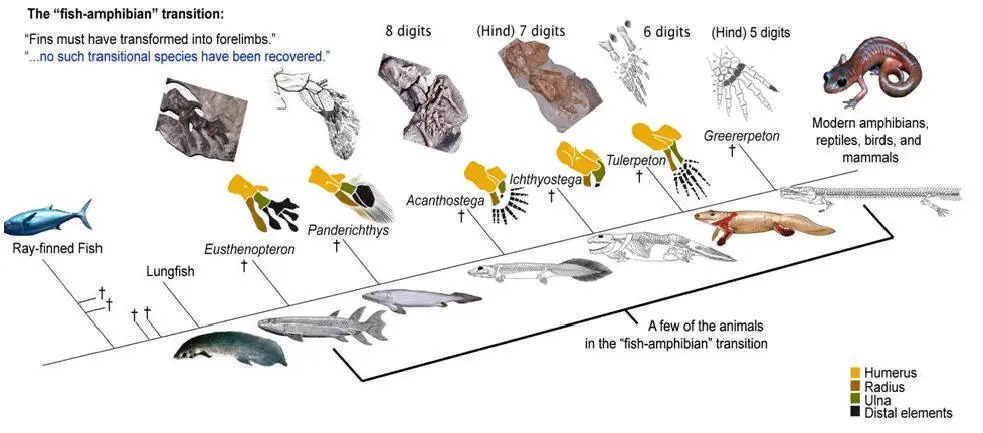
Упрощенная схема перехода от лопастеперых рыб к первым тетраподам. Этот рисунок использовался на «обезьяньем процессе» в Пенсильвании в 2005 г.для иллюстрации ложности заявлений креационистов об отсутствии переходных форм между рыбами и амфибиями (процесс, как известно, выиграли эволюционисты, несмотря на то, что судья Джонс, выносивший вердикт, был консерватором, сторонником Буша и очень религиозным человеком). На этой картинке еще нет тиктаалика, который был найден позже.

Более точные рисунки передних конечностей пандерихта и тиктаалика (предоставил А.Н.Кузнецов).
Уже у Eusthenopteron в переднем плавнике имелась кость, соответствующая плечевой кости тетрапод (показана желтым цветом), и две кости, соответствующие будущим локтевой и лучевой костям (показаны коричневым и зеленым цветом). «Дистальные элементы», соответствующие будущей кисти тетрапод, показаны черным цветом. У пандерихта строение этих дистальных элементов еще оставалось относительно неупорядоченным. У тиктаалика они приобрели чуть более «оформленный» вид, так что становится уже более-менее понятно, откуда взялись у потомков тиктаалика пальцы. Тиктаалик и пандерихт уже могли сгибать свой передний то ли плавник, то ли лапу в суставе, который у их потомков назовут «локтевым». Также у тиктаалика произошла утрата жаберной крышки (которая еще была у пандерихта), что еще на шажок приближает тиктаалика к тетраподам. Утрата жаберной крышки автоматически привела к исчезновению костной связи между поясом передних конечностей и черепом.
У вентастеги и ряда других форм, которые по строению черепа занимают промежуточное положение между тиктааликом и акантостегой, к сожалению, от конечностей почти ничего не сохранилось.
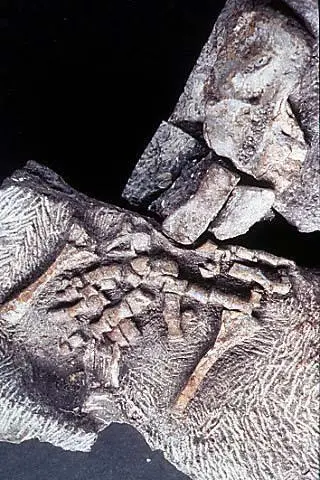
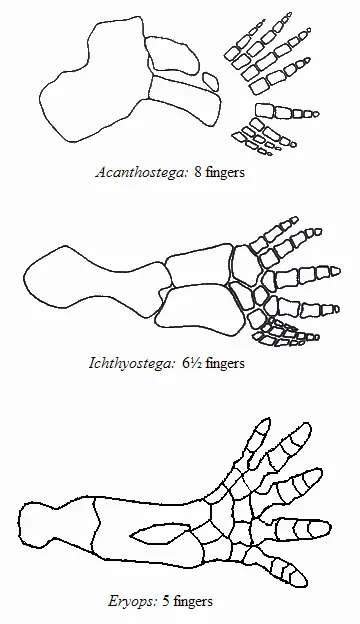
Передняя конечность акантостеги и других ранних тетрапод
Конечности древнейших животных, которых безоговорочно относят уже к тетраподам, то есть к первым амфибиям (акатностега, ихтиостега), представляют собой всего лишь еще один небольшой шажок в сторону дифференциации дистальных частей скелета, т. е. кисти. У акантостеги еще сохранялись внутренние жабры, у ихтиостеги они уже редуцировались (наружные жабры у личинок амфибий сохраняются и сегодня). В дальнейшем у древнейших амфибий происходило постепенное развитие шейного отдела. Совершенно непонятно, где в этом ряду находится искомый великий «макропереход». Сказать, что переход от эустеноптерона к пандерихту и от пандерихта к тиктаалику — это «микро», а вот от тиктаалика к акантостеге — это уже «макро», может только очень предвзятый человек. И, конечно, среди специалистов-палеонтологов нет никого, кому пришло бы в голову это утверждать.
Читать дальшеИнтервал:
Закладка:



