М. Канунго - Биохимия старения

- Название:Биохимия старения
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:1982
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
М. Канунго - Биохимия старения краткое содержание
Предназначена для биологов, биохимиков, геронтологов, врачей-гериатров.
Биохимия старения - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
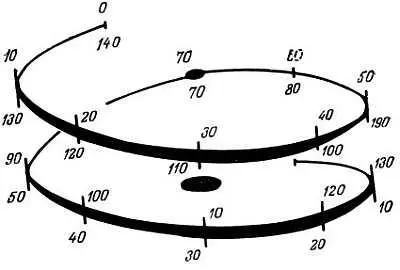
Рис. 2.3. Предполагаемое закручивание суперспирали ДНК вокруг сердцевины нуклеосомы. Отмечены места расщепления ДНК нуклеазой [117]
Сердцевина нуклеосомы содержит по две молекулы каждого из Н2А-, Н2В-, Н3- и Н4-гистонов, которые образуют октамер. Положительно заряженные вытянутые цепи этих гистонов электростатически связаны с отрицательно заряженной ДНК. Полагают, что четыре гистона расположены относительно ДНК следующим образом:
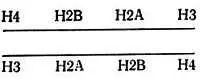
Два гистона, Н3 и Н4, богатые аргинином, вероятно, взаимодействуют с двумя концами фрагмента ДНК. Когда эти гистоны добавляют к двухцепочечной ДНК, они образуют характерную структуру типа бублика, видимую в электронный микроскоп [129]. При воссоединении гистонов сердцевины со 140 парами оснований ДНК образуются частицы, имеющие тот же самый коэффициент седиментации, что и нуклеосомы, полученные из хроматина [36, 345]. Было также показано, что одни гистоны Н3 и Н4 образуют с ДНК структуры, похожие на сердцевины нуклеосом, устойчивые к трипсину [64, 327] и дающие картину дифракции рентгеновских лучей, похожую на картину для нативных нуклеосом [261]. Когда гистоны Н3 и Н4 добавляют к ДНК, они связываются со 140 парами оснований ДНК, которая имеет 1,5 сверхспиральных оборота вокруг тетрамера [195]. Образующаяся структура представляет собой цилиндр с размерами 45×8×8 нм. При последующем добавлении гистонов Н2А и Н2В цилиндр сжимается и становится похожим на нативную нуклеосому. Аналогичные явления наблюдал Картер [70]. Это согласуется с высказанным ранее [198] предположением, что гистоны Н3 и Н4 играют существенную роль в образовании структуры нуклеосомы. Эти два гистона наиболее консервативны, содержат большое количество β-структур и взаимодействуют друг с другом сильнее, чем с другими гистонами. По степени связывания с ДНК гистоны располагаются в следующем порядке: Н3 и Н4>Н2А>Н2В>Н1 [283]. При изучении поперечных сшивок показано, что связаны следующие пары: Н3-Н4, Н2А-Н2В и Н2В-Н4 [84].
Согласно одной из точек зрения, сначала 2 молекулы гистона Н3 и 2 молекулы гистона Н4 образуют тетрамер и связываются со 140 парами оснований ДНК, формируя основную сердцевину нуклеосомы. На втором этапе в эту структуру включаются по две молекулы гистонов Н2А и Н2В, чем и завершается образование нуклеосомы [42, 64, 258, 372]. При изучении сборки новореплицированного хроматина Drosophila показано, что гистоны Н3 и Н4 соединяются с ДНК в течение или вскоре после ее синтеза, гистоны Н2А и Н2В — на 2-10 мин позже, а гистон Н1 — через 10–20 мин, и в результате образуется зрелый хроматин [375]. По-видимому, во взаимодействие с ДНК вовлечены COOH-концы четырех гистонов, так как удаление ЫН2-концевых участков цепей гистонов не влияет на структуру нуклеосомы [371]. Гистоны Н2А и Н2В образуют димеры, взаимодействуя своими центральными неполярными областями, так что NH 2- и COOH-концы остаются свободными. Гистоны Н3 и Н4 образуют димеры путем образования связей между их центральными неполярными областями и COOH-концами, так что основные NH 2-концевые области нуклеосомных гистонов доступны для взаимодействия с кислотными группами ДНК [72]. Роль NH 2-концевых областей четырех гистонов пока не установлена, хотя известно, что они связываются с ДНК. Мирзабеков и др. [252] путем ковалентных сшивок гистонов с 5′-концевыми фрагментами ДНК показали, что каждый гистон связан с 10 парами оснований ДНК. Сборка нуклеосом, по-видимому, контролируется НГБ. Так, очищенный препарат этих белков, выделенный из яиц Xenopus laevis , в бесклеточной системе в присутствии гистонов и очищенной ДНК катализирует образование нуклеосом [217].
Таким образом, основная структура хроматина представляет собой цепь линейно расположенных нуклеосом диаметром 10 нм, которую называют нуклеосомной фибриллой. Это низший уровень организации хроматина. Структуру более высокого порядка образуют нуклеосомы, свернутые в спираль, которая имеет диаметр 20–30 нм и шаг 10 им. Свертывание нуклеосом в спираль, по-видимому, обеспечивается богатым лизином гистоном Н1, который, как было показано, соединяется с линкерной ДНК между соседними нуклеосомами. Этот вывод следует из того, что после расщепления мононуклеосом стафилококковой нуклеазой размер ДНК уменьшается с 200 до 140 пар оснований, причем одновременно освобождается 35-парный фрагмент ДНК, связанный с гистоном Н1 [20]. Когда гистон Н1 добавляли к хроматину, который был его лишен, увеличение сродства к нему наблюдалось только до стадии образования октануклеосомы, но не далее [301]. Связывание с гистоном Н1 не только стабилизирует ДНК в линкерной области, но вызывает также ее дальнейшую конденсацию и свертывание [75]. Более высокий порядок структуры хроматина (по сравнению с цепочкой бусин) представляет собой спираль из частиц октануклеосом, образование которой обеспечивается гистоном Н1 или гистоном Н5 (в случае эритроцитов, содержащих ядра). Это согласуется с результатами, согласно которым полинуклеосомы, содержащие около шести нуклеосом, являются, по-видимому, основными матрично активными единицами хроматина, связывающимися с эндогенной РНК-полимеразой [344]. Олигонуклеосомы служат лучшими матрицами для транскрипции, чем мононуклеосомы, и на них синтезируются более длинные транскрипты [318].
Нуклеосома — динамическая единица как в структурном, так и в функциональном отношении. Как сказано выше, она состоит из двух половин, что может быть определено путем специфического связывания восьми молекул гистонов с ДНК. То, что нуклеосомы в транскрипционно активном состоянии подвержены конформационным изменениям, становится очевидным при изучении их чувствительности к ДНКазе I. Этот фермент преимущественно воздействует на те последовательности ДНК, которые активно транскрибируются. Он удаляет ДНК, кодирующую глобин, из ядер эритроцитов цыпленка, но не действует на ядра клеток мозга или фибробластов [125, 282, 367]. На ДНК яичного альбумина эритроцитов и фибробластов, в которой этот ген не транскрибируется, фермент также не действует. Стафилококковая нуклеаза, которая, как известно, расщепляет ДНК в межнуклеосомной области, не расщепляет ДНК глобина из эритроцитов цыпленка. Если мономерные нуклеосомы, полученные из этих клеток действием стафилококковой ДНКазы, обработать затем ДНКазой I, то преимущественно удаляются гены глобина. Показано [125], что ген яичного альбумина предпочтительно расщепляется ДНКазой в клетках яйцевода курицы и не расщепляется в других клетках, в которых он не транскрибируется. В клетках хомяка, трансформированных аденовирусом, последовательности ДНК аденовируса, которые легко расщепляются ДНКазой I, представляют собой участки, с которых транскрибируется мРНК. Другие вирусные последовательности резистентны к этой нуклеазе [119]. Из приведенных наблюдений следует, что во время транскрипции происходят конформационные изменения в хроматине, так что ДНК становится более чувствительной к ДНКазе I, но ее чувствительность к стафилококковой нуклеазе остается прежней. Полученные результаты подтверждаются данными электронной микроскопии [313]. Показано, что в процессе развития ооцитов трех видов Xenopus транскрипционно активный ядрышковый хроматин выглядит гладким, нуклеосомы в нем присутствуют в небольшом количестве или вообще отсутствуют. Неактивный хроматин имеет вид бусин. Пониженная транскрипционная активность хроматина коррелирует с появлением бусин в его структуре, тогда как транскрипционно активный хроматин содержит больше мононуклеосом, чем транскрипционно неактивный, что и означает увеличение той области хроматина, которая активна при транскрипции [223]. Электронно-микроскопическое изучение активно транскрибируемых рибосомных генов Physarum polycephalum показывает, что ДНК в транскрибируемом участке имеет вытянутую конформацию [179]. Таким образом, структура хроматина и, в особенности, нуклеосом подвержена конформационным изменениям в процессе транскрипции, а возможно, и репликации. Не исключено, что это вызвано связыванием с НГБ. Для ковалентной модификации гистонов различных типов, па-пример фосфорилирования, ацетилирования, метилирования и ADPрибозилирования, необходимы эффекторы.
Читать дальшеИнтервал:
Закладка:









