М. Канунго - Биохимия старения

- Название:Биохимия старения
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:1982
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
М. Канунго - Биохимия старения краткое содержание
Предназначена для биологов, биохимиков, геронтологов, врачей-гериатров.
Биохимия старения - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Некоторые исследователи изучали содержание и сродство мембранных рецепторов к соответствующим гормонам как функцию возраста. Активность аденилатциклазы, стимулированная адреналином (но не глюкагоном), в мембране клеток печени крыс в ходе развития уменьшается [11]. В адипоцитах уменьшается активность аденилатциклазы, чувствительной к глюкагону [34]. В коре головного мозга наблюдается падение с возрастом определяемого норадреналином содержания сАМР [10]. Это проявляется в уменьшении активности аденилатциклазы [62]. Кроме того, с возрастом уменьшается чувствительность клеток коры головного мозга к норадреналину и дофамину [43, 62]. Адренергическое связывание в клетках мозжечка крыс и человека с возрастом также снижается [33]. Таким образом, дисфункция мозжечка в старческом возрасте может быть обусловлена утрачиванием нейронов, получающих норадренергический импульс. У старых людей концентрация β-адренергических мембранных рецепторов в лейкоцитах значительно ниже, чем в среднем возрасте, хотя сродство рецепторов к (-) 3Н-дигидроальпренелолу с возрастом не меняется [57]. Было обнаружено [18] дифференцированное (падение при старении активности аденилатциклазы, стимулированной норадреналином, глюкагоном и АКТГ. Неизвестно, вызвано ли это изменением рецепторов гормонов или самой аденилатциклазы. Связывание (-) 3Н-дигидроальпренелола и гамма-ашшомасляной кислоты с синаптическими мембранами в некоторых участках мозга с возрастом уменьшается [42]. Связывание 3Н-атропина с синаптосомами из коры головного мозга старых крыс значительно ниже, чем у более молодых животных (рис. 5.9) [28]. Следовательно, по окончании репродуктивного периода уменьшается содержание в мозгу мускариновых рецепторов ацетилхолина.
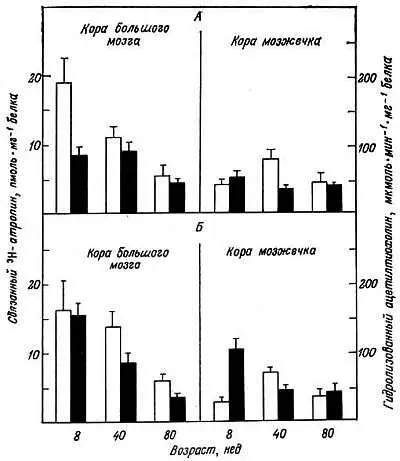
Рис. 5.9. Специфическое связывание 3Н-атропина с синаптосомальными фракциями (светлые столбики) и гидролиз ацетилтиохолина (темные столбики) в коре большого мозга и коре мозжечка у крыс-самок (А) и самцов (Б) разного возраста [28]
Приведенные выше данные, очевидно, достаточно сложны, так как они отражают различные взаимодействующие процессы. Кроме того, как и в случае ферментов, разные исследователи выражали содержание рецепторов разными способами. Уровень рецептора может зависеть от физиологических условий эксперимента. Некоторые использовали для определения содержания рецепторов гомогенат целиком, тогда как другие — его отдельные фракции. Оптимальная активность рецептора может быть обусловлена присутствием некоторых факторов, свойственных цитозолю, которые могут отсутствовать в отдельных фракциях. Кроме того, по всей вероятности, с возрастом меняется структура самих мембран, что влияет на центры взаимодействия с гормонами в молекулах рецептора.
Среди результатов, связанных с изменениями мембранных рецепторов, наблюдались, однако, некоторые исключения. Так, содержание рецепторов инсулина в печени [25], β-адренергических рецепторов в эритроцитах [13], рецепторов глюкагона в адипоцитах уменьшается в ранний период развития, а не по окончании репродуктивного периода. Очевидно, поддержание уровня этих рецепторов связано с определенными функциями, и эти функции не нарушены. Следует напомнить, что для начала процесса старения не обязательно ухудшение или нарушение всех функций организма. Кроме того, изменение содержания рецепторов одних и тех же гормонов может быть различным в премитотических и постмитотических клетках. Разные клетки и ткани, по-видимому, начинают стареть в разное время, и скорости их старения с точки зрения изменения специфических функций могут быть различными.
Большое значение имеет тот факт, что уровень рецептора регулируется не только соответствующим гормоном, но также и другими гормонами и эффекторами. Показано [40], что при введении крольчихам 17β-эстрадиола увеличиваются концентрация рецептора окситоцина и сократимость матки. Вместе с тем прогестерон уменьшает уровень рецептора окситоцина и сократимость матки. Таким образом, содержание одного гормона может зависеть от других гормонов или эффекторов.
Степень ответа клеток или тканей-мишеней на Действие гормонов является наилучшим показателем активности последних. В табл. 5.1 этот показатель для отдельных гормонов рассмотрен как функция возраста. Оптимальный ответ на действие гормона зависит от нескольких факторов, а именно: содержания гормона (которое можно варьировать путем экзогенного введения), содержания рецептора, проникновения гормона в ядро в случае стероидов, активации аденилатциклазы в клеточной мембране в случае пептидных и других гормонов, связывания комплекса Г-Р с хроматином и т. д. Одним из лучших тестов на силу ответа ткани на действие гормона является индукция какого-либо фермента. В ряде работ этот метод использован при введении in vivo и in vitro стероидных гормонов [1, 2, 24, 31]. Было получено четыре варианта изменения индукции [2]: а) в старческом возрасте увеличивался лаг-период, но не менялась величина индукции; 6) уменьшалась или увеличивалась величина индукции; в) менялись и лаг-период, и величина индукции; г) оба параметра оставались постоянными (гл. 3). Было обнаружено также, что индукция стероидными гормонами малатдегидрогеназы, ацетилхолинэстеразы, глутаминсинтетазы, аргиназы, аланинаминотрансферазы и тирозинаминотрансферазы ингибируется актиномицином D [31, 39, 41, 44–46]. Этот факт указывает на то, что индукция в данном случае происходит путем стимуляции транскрипции соответствующих мРНК. Индукция большинства этих ферментов в старческом возрасте нарушается, и это нарушение может происходить на уровне либо транскрипции, либо экспрессии генов, соответствующих изучаемым ферментам. Кроме того, как было показано на примере пируваткиназы, возрастные изменения индукции ферментов зависят также от пола животных и от типа ткани [15–17].
Ответ ткани на действие специфических гормонов может модулироваться другими гормонами. Было показано, что после обработки эксплантатов молочной железы крысы пролактином синтезируется мРНК казеина ([35]. Если в инкубационную среду одновременно с пролактином добавляют гидрокортизон, то синтез мРНК значительно возрастает. Однако если вместо гидрокортизона добавляют прогестерон, то синтез мРНК подавляется. Действие пролактина осуществляется с помощью мембранного рецептора, тогда как действие прогестерона и гидрокортизона — с помощью рецепторов, имеющихся в цитозоле. Как именно пролактин стимулирует синтез мРНК казеина, точно неизвестно, но эти данные представляют собой прекрасный пример того, как разные гормоны, имеющие разные механизмы действия, модулируют активность одного и того же гена. Еще в одной работе, в которой использовали культуру клеток гипофиза, было показано, что секреция этими клетками пролактина стимулируется 17β-эстрадиолом, а этот эффект ингибируется дофамином — основным ингибирующим веществом гипоталамуса [48]. 17β-эстрадиол снимает ингибирующий эффект дофамина, возможно действуя на его рецепторы.
Читать дальшеИнтервал:
Закладка:









