М. Канунго - Биохимия старения

- Название:Биохимия старения
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:1982
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
М. Канунго - Биохимия старения краткое содержание
Предназначена для биологов, биохимиков, геронтологов, врачей-гериатров.
Биохимия старения - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
21. Harman D . Radiat. Res., 16, 753–763 (1962).
22. Harman D . J. Gerontol., 23, 476–482 (1968).
23. Hasan M., Glees P . Expl. Gerontol., 7, 345–351 (1972).
24. Hasan M., Glees P . Expl. Gerontol., 8, 78–83 (1973).
25. Hasan M., Glees P., Spoerri P. E . Cell Tiss. Res., 150, 369–375 (1974).
26. Hochschild R . Expl. Gerontol., 8, 117–183 (1973).
27. Hodge C. F . J. Physiol., 17, 129–134 (1894).
28. Kormendy C. G., Bender A. D . Gerontologia, 77, 52–64 (1971).
29. Miquel J., Lundgren P. R., Johnson J. E . J. Gerontol., 33, 5-19 (1978).
30. Munkres K. D., Rana R. S . Mech. Age. Dev., 7, 399–406 (1978).
31. Munkres K. D., Rana R. S . Mech. Age. Dev., 7, 407–415 (1978).
32. Nandy K., Bourne G. H . Nature, 210, 313–314 (1966).
33. Nandy K . Mech. Age. Dev., 8, 131–138 (1978).
34. Nandy K. J . Am. Gerontol. Soc, 26, 74–81 (1978).
35. Pryor W. A . Sci. Amer, 23, 70–83 (1970).
36. Pryor W. A . Fed. Proc, 32, 1862–1869 (1973).
37. Riga S., Riga D . Brain Res., 72, 265–275 (1974).
38. Samorajski T., Keefe J. R., Ordy J. M . J. Gerontol., 19, 262–276 (1964).
39. Shimasaki H., Nozawa T., Privett O. S., Anderson W. R . Arch. Biochem. Biophys., 183, 443–451 (1977).
40. Sinex F. M . In: The Handbook of the Biology of Aging (C. E. Finch and L. Hayflick, Eds.), 43–46, Reinhold, New York (1977).
41. Spence A. M., Herman N. M . Mech. Age. Dev., 2, 211–227 (1973).
42. Spoerri P. E., Glees P . Expl. Gerontol., 8, 259–263 (1973).
43. Spoerri P. E., Glees P . Mech. Age. Dev., 3, 131–155 (1974).
44. Strehler B. L . Adv. Gerontol. Res., 1, 343–384 (1964).
45. Strehler B. L., Mark D. D., Mildvan A. S., Gee M. V . J. Gerontol., 14, 430–439 (1951).
46. Sulkin N. M., Srivanji P. J . Gerontol, 15, 2–9 (1960).
47. Szabo D., Desinick C., Okros L., Stack E . Expl. Gerontol., 5, 335–337 (1970).
48. Tappet A. L . Ann. N. Y. Acad. Sci., 203, 12–28 (1972).
49. Tonna E. A . Expl. Gerontol., 8, 9-16 (1973).
50. Tonna E. A . J. Gerontol., 30, 3–8 (1975).
51. Treff W. M . Altern, 37–54, Schattauer, Stuttgart (1974).
52. Verzar F . Gerontologia, 4, 104–111 (1960).
53. Verzar F . Sci. Amer., 208, 104–114 (1963).
54. Verzar F . Intern. Rev. Connect. Tissue Res., 2, 243–299 (1964).
55. Zeman W . Adv. Gerontol. Res., 3, 147–170 (1971).
Глава 7. Изменения иммунной системы
Животные к старости становятся более восприимчивыми к заболеваниям. За защиту организма от болезней, которые вызываются патогенными бактериями, вирусами, грибами и другими чужеродными агентами, ответственна иммунная система. Эта система не очень хорошо развита у беспозвоночных, но у позвоночных она претерпела значительную эволюцию [127]. Птицы и млекопитающие обладают высокоразвитой и сложной иммунной системой. Млекопитающее способно защитить себя практически от любого чужеродного, "не своего" вещества, которое проникает в его организм. Такие вещества, которые не представлены в организме, называют антигенами. Иммунная система ответственна за сохранность любого органа, который может быть поврежден антигеном. Следовательно, нарушения функций иммунной системы при старении делают организм более восприимчивым к патогенным факторам, увеличивают частоту заболеваний и, следовательно, снижают функциональную активность организма. Имеется несколько основательных обзоров, касающихся иммунной системы и ее возрастных изменений [19, 24, 25, 48, 58, 70].
Иммунная система позвоночных состоит из костного мозга, тимуса, лимфатических узлов и селезенки (рис. 7.1). У птиц важной частью этой системы является фабрициева сумка. Она представляет собой дивертикул клоаки и является по природе лимфоидным образованием. Клетками, ответственными за создание иммунитета или за предохранение животного от антигена, являются лимфоциты. Развитие иммунокомпетентных лимфоцитов происходит у птиц и млекопитающих следующим образом [19]. Стволовые клетки лимфоцитов происходят из желточного мешка эмбриона. Они мигрируют в печень и селезенку эмбриона и далее в костный мозг. Определенные предшественники лимфоцитов мигрируют из костного мозга в тимус, где они, вероятно, подвергаются некоторой дифференцировке и формируются. Эти клетки затем покидают тимус и расселяются в тимус-зависимой зоне лимфатических узлов и селезенки. Поскольку они формируются в тимусе, их называют Т-лимфоцитами.
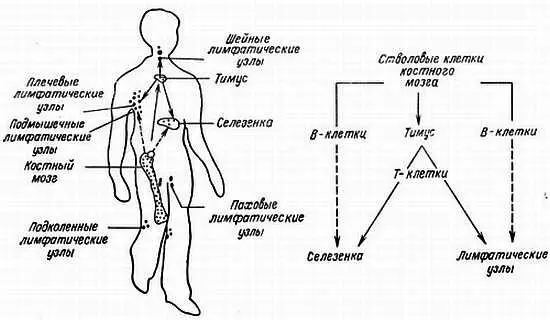
Рис. 7.1. Формирование клеток иммунной системы [70]
Т-клетки ответственны за клеточные иммунные реакции, которые защищают организм от патогенных грибов и вирусов. Они также ответственны за отторжение чужеродных тканей и трансплантатов — пересаженных органов и опухолей, за повышенную чувствительность (аллергические реакции) к некоторым антигенам и за аутоиммунные реакции. Иммунный ответ с участием Т-клеток характеризуется непосредственной атакой этими клетками антигена, поэтому такой иммунный ответ называют клеточным ответом, или клеточным иммунитетом. Различают следующие три класса Т-клеток: стимуляторы (хелперы), супрессоры и киллеры. Полагают, что они дифференцируются в тимусе таким образом, что приобретают специфические рецепторы на своей поверхности для распознавания специфических антигенов. Т-хелперы кооперируются с антигенспецифическими В-лимфоцитами и стимулируют выработку специфических белков, обозначаемых в дальнейшем антителами. Супрессорные Т-клетки подавляют выработку антител специфическими В-клетками к специфическим антигенам и, кроме того, препятствуют выработке В-клетками антител к клеткам и молекулам собственного организма. Специфические Т-киллеры распознают антигены типа трансплантатов от несовместимых доноров, опухолей, грибов и т. д., взаимодействуют с ними с помощью своих рецепторов и инактивируют или нейтрализуют их. Функции Т-клеток, возможно, реализуются через секреторные факторы, называемые лимфокинами, которые еще не полностью охарактеризованы. Т-клетки можно отделить от В-клеток, так как они имеют различные иммуноглобулиноподобные поверхностные рецепторы [113, 127]. Если удалить тимус у только что вылупившихся цыплят или новорожденных мышей и крыс, то клеточный иммунитет нарушается. Трансплантация тимуса молодых мышей старым особям или мышам, облученным с целью разрушения их иммунокомпетентных клеток, восстанавливает клеточный иммунитет у реципиентов. Таким образом, тимус участвует в клеточном иммунитете.
У птиц определенные лимфоциты мигрируют из костного мозга в фабрициеву сумку и спустя некоторое время — в лимфатические узлы и селезенку. Эти клетки называются В-лимфоцитами, поскольку они дифференцируются и созревают в фабрициевой сумке (англ. bursa). Эквивалент фабрициевой сумки у млекопитающих не известен. Предполагают, что местом созревания В-лимфоцитов может быть костный мозг. В-клетки ответственны за гуморальный иммунитет. При этом виде ответа клон В-клеток в лимфатических узлах и селезенке после контакта со специфическим антигеном, который достигает их по кровотоку, делится 6–8 раз для выработки высокодифференцированных плазматических клеток. Антиген каждого типа распознается специализированной В-клеткой только одного типа или клоном В-клеток. Для трансформации В-клеток в плазматические требуется их кооперация со специфическими Т-хелперами, также присутствующими в лимфоидных органах. Образующиеся плазматические клетки синтезируют большое число специфических белковых молекул, называемых антителами, или иммуноглобулинами (Ig), которые выделяются в циркулирующую жидкость, гумор. Иммуноглобулины циркулируют в жидкостях организма и при встрече с антигеном, индуцировавшим их синтез, связываются с ним и нейтрализуют его. Млекопитающие и птицы способны вырабатывать иммуноглобулины практически к любому антигену. Иммуноглобулины ответственны за иммунитет к бактериям и вирусам. Поскольку активное начало этого типа иммунитета присутствует в жидкости (гуморе), он называется гуморальным иммунитетом.
Читать дальшеИнтервал:
Закладка:









