М. Канунго - Биохимия старения

- Название:Биохимия старения
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:1982
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
М. Канунго - Биохимия старения краткое содержание
Предназначена для биологов, биохимиков, геронтологов, врачей-гериатров.
Биохимия старения - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
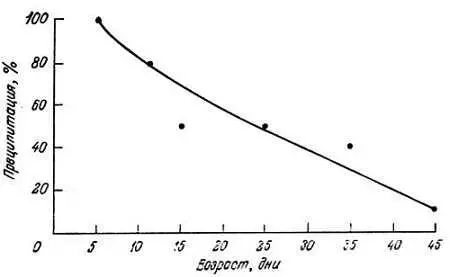
Рис. 9.4. Зависимость общей активности изоцитрат-лиазы, преципитированной данным количеством антител, от возраста круглого червя Tyrbatrix aceti [100]
Были сделаны попытки определить накопление ошибок в белках в зависимости от возраста культивируемых фибробластов. Холлидей и Террент [49] описали увеличение доли неактивной и термолабильной глюкозо-6-фосфат — дегидрогеназы на поздних пассажах фибробластов MRC-5. Лактатдегидрогеназа фибробластов поздних пассажей этой линии отличалась по антигенности от фермента ранних пассажных клеток [71]. Голдстайн и Мёрмен [37] представили сходные данные по трем ферментам фибробластов кожи человека. Имеются данные не только о значительном уменьшении точности работы ДНК-полимеразы в клетках поздних пассажей (что изучалось с помощью синтетических матриц), но и об уменьшении скорости элонгации репликона [73]. Большой процент ошибок, регистрируемый этими авторами, вряд ли присущ нормальным клеткам; поскольку он был бы для них катастрофичным, не исключено, что регистрируемая неточность считывания обусловлена использованием в качестве матриц синтетических полинуклеотидов. Мартин и др. [79] не обнаружили различий в термочувствительности и антигенности ферментов клеток ранних и поздних пассажей в культуре WI-38. Следовательно, иммунные и каталитические свойства, а также положение при электрофокусировании фосфоглицераткиназы, пируваткиназы типа М 2, глюкозофосфатизомеразы и глюкозо-6-фосфат — дегидрогеназы не отличаются у фибробластов ранних и поздних пассажей. Не изменяется также скорость, а следовательно, и надежность репликации вирусов.
Ферменты β-N-ацетилглюкозаминидаза, α-глкжозидаза и α-N-ацетилгалактозаминидаза лизосом и NADH-дегидрогеназа митохондрий молодых и старых фибробластов WI-38 имеют сходную термолабильность [52]. РНК-полимераза, которая участвует в синтезе белка и, согласно Оргелу, является одним из основных виновников появления ошибок, была изучена Ивенсом [25]. Он не нашел различий в термолабильности и удельной активности ферментов, полученных из молодых и старых клеток.
Гервин и др. [29] измерили ошибки считывания poly (U) in vitro в неочищенных препаратах, полученных в различное время из E. coli , растущей в присутствии дигидрострептомицина, который, как известно, вызывает ошибки трансляции. Они наблюдали значительное увеличение ошибок считывания poly (U), но их уровень достигал стабильных значений в присутствии антибиотика в течение нескольких генераций. Исследования Эделмена и Гелланта [24] in vivo дали сходные результаты. Оценка ошибочной трансляции (включение цистеина в флагеллин) у E. coli в присутствии стрептомицина показала, что частота ошибки становится в 50 раз выше, чем в норме, и затем стабилизируется на одном уровне (рис. 9.5). Таким образом, даже если ошибки появляются в процессе трансляции, то, как и предполагали Гоел и Икес [35], достигается стабилизация ее точности. Поэтому маловероятно, чтобы первопричиной старения было возникновение ошибок в процессе переноса информации.
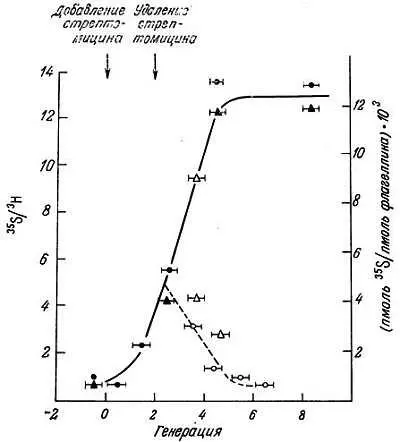
Рис. 9.5. Кинетика частоты ошибки в присутствии и в отсутствие стрептомицина [24]. Культура АС92 была выращена в отсутствие стрептомицина, и в ней была измерена частота фоновой ошибки. В момент, отмеченный стрелкой, добавляли 5 мкг/мл стрептомицина и измеряли частоту ошибки на протяжении более чем восьми последующих генераций. После двух генераций в присутствии стрептомицина из культуры отбирали аликвоту, отмывали минимальной средой без добавок для удаления стрептомицина и ресуспендировали в среде с добавками. Снова вычисляли частоту ошибки. Культуру метили 3Н-аланином (53 Ки/ммоль) и 35S-сульфатом (2 мКд/нмоль) и выделяли флагеллин, как описано ранее [24]. Сплошная линия и темные значки — частота ошибок в клетках, растущих в присутствии стрептомицина; штриховая линия и светлые значки — частота ошибок в клетках после удаления стрептомицина; светлые и темные кружки и треугольники обозначают данные, полученные в двух экспериментах. Длина горизонтальных линий на значках указывает на фракции той генерации, в пределах которой включалась метка
В заключение можно сказать, что существует очень много данных о снижении в пожилом возрасте на 30–70 % удельной активности альдолазы А и В [32, 33] и пероксид-дисмутазы у крыс [[98, 99], а также изоцитрат-лиазы [31, 100], фруктозобисфосфат-альдолазы [133], енолазы [111] и фосфоглицераткиназы [41] у Turbatrix aceti . Для нескольких ферментов установлены различия в термолабильности и антигенных свойствах. Вместе с тем удельная активность триозофосфатизомеразы Turbatrix aceti [40] и орнитиндекарбоксилазы печени крыс [86, 87] не изменяется.
При измерении других параметров перечисленных ферментов: К м, K i, электрофоретической подвижности и молекулярной массы никаких существенных различий не обнаружено. Каковы же причины возрастных изменений свойств ферментов? Использование совершенной методики типа изоэлектрофокусирования, с помощью которой можно разделять белки, отличающиеся только суммарным электрическим зарядом, не выявило никаких отличий между пероксид-дисмутазами молодых и старых крыс [39, 98, 99]. Более того, когда провели двумерное картирование белков верхних шейных нервных узлов молодых и старых крыс, также не нашли никаких различий, причем в одном направлении белки разделялись в соответствии с их изоэлектрической точкой, а в другом — в соответствии с их молекулярной массой [129]. Триптические карты актина и миозина скелетной мышцы молодых и старых крыс оказались одинаковыми [117].
Таким образом, имеются убедительные данные, которые показывают, что с увеличением возраста ошибки типа замещения аминокислот в белках не возникают в сколько-нибудь существенном количестве ни in vivo, ни in vitro. Появление ошибок является случайным процессом, и если бы они возникали, то существовали бы белки с замещенными аминокислотами, однако такие белки не обнаружены. Кроме того, более или менее постоянную продолжительность жизни видов и постепенное ослабление функции с увеличением возраста нельзя объяснить теорией ошибок, так как для того, чтобы иметь отношение к упомянутым феноменам, ошибки должны возникать с определенной скоростью. Приходится предполагать, что частота ошибок регулируется генами или другими факторами, что противоречит основной идее теории ошибок. Следовательно, маловероятно, чтобы причиной старения было нарастание ошибок в функциональных макромолекулах с возрастом.
Читать дальшеИнтервал:
Закладка:









