Александр Марков - Эволюция. Классические идеи в свете новых открытий

- Название:Эволюция. Классические идеи в свете новых открытий
- Автор:
- Жанр:
- Издательство:АСТ: CORPUS
- Год:2014
- Город:Москва
- ISBN:978-5-17-083218-7
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Александр Марков - Эволюция. Классические идеи в свете новых открытий краткое содержание
Книга Александра Маркова и Елены Наймарк рассказывает о новейших исследованиях молекулярных генетиков и находках палеонтологов, которые дают ответы на эти и многие другие вопросы о видоизменениях в природе. Тысячи открытий, совершенных со времен Дарвина, подтверждают догадки родоначальников теории эволюции; новые данные ничуть не разрушают основы эволюционной теории, а напротив, лишь укрепляют их.
Александр Марков, заведующий кафедрой биологической эволюции биофака МГУ, и Елена Наймарк, ведущий научный сотрудник Палеонтологического института им. А. А. Борисяка, — известные ученые и популяризаторы науки. Двухтомник «Эволюция человека» (2011), написанный ими в соавторстве, стал настольной книгой не только для студентов и ученых-биологов, но и для множества людей за пределами профессионального сообщества.
Эволюция. Классические идеи в свете новых открытий - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Поскольку для крупных эволюционных преобразований требуется последовательное закрепление множества мутаций, такие изменения трудно «расшифровать» на молекулярном уровне, а уж пронаблюдать воочию и вовсе невозможно из-за длительности процесса. Те случаи, которые все-таки удается расшифровать, — это, по-видимому, редкие, нетипичные случаи, когда крупное изменение обеспечивается всего несколькими мутациями. Но такие случаи есть, и мы пока не знаем наверняка, так ли уж мала их роль на больших — эволюционно значимых — отрезках времени. Мы познакомимся с одним из таких случаев, когда всего две мутации обеспечили интересное и важное адаптивное приобретение у многоклеточного животного — нематоды (круглого червя). В результате двух мутаций нематоды приобрели способность к самооплодотворению — стали гермафродитами. Этот пример важен нам также для того, чтобы показать, как в ходе эволюции подстраиваются друг к другу разные гены, регулирующие определенную функцию.
У большинства нематод, как и у многих других животных, пол определяется генетически, при помощи половых хромосом. Если в оплодотворенном яйце две X-хромосомы, из яйца разовьется самка, если одна — самец ( Haag, 2005 ). Однако у двух наиболее изученных видов нематод, Caenorhabditis elegans и C. briggsae , особи с двумя X-хромосомами — не самки, а гермафродиты. Их гонады (половые железы) на поздних личиночных стадиях производят спермии. Нематоды утратили в ходе эволюции жгутики, поэтому и спермии у них безжгутиковые. Они похожи на амеб и передвигаются при помощи псевдоподий. У гермафродитов безжгутиковые спермии поздних личинок переползают в специальные хранилища — сперматеки — и здесь ждут своего часа. Гонады взрослых гермафродитов производят уже не спермии, а яйцеклетки. Они могут быть оплодотворены как собственными спермиями из сперматеки, так и спермиями самца в результате спаривания.
Предки C. elegans и C. briggsae были нормальными раздельнополыми червями. Это следует, в частности, из того, что все прочие виды рода Caenorhabditis — раздельнополые. По-видимому, гермафродитизм у C. elegans и C. briggsae является «эволюционно молодым», новым признаком. Чтобы разобраться, как он возник, понадобились выдумка, знание геномов нескольких видов нематод и эксперименты, осуществленные американскими биологами ( Baldi et al., 2009 ). Вот как это было.
Для начала уточним, какие гены работают при формировании половых различий у нематод. Наличие одной (а не двух) X-хромосом у самцов служит триггером, включающим синтез белка HER-1, который ингибирует белок TRA-2. Подавление активности TRA-2 через пару промежуточных шагов приводит к уничтожению белка TRA-1, функция которого состоит в том, чтобы отключать ряд ключевых генов, направляющих развитие по «мужскому» пути (в том числе ген fog-3 , контролирующий сперматогенез) [29] Названия генов принято писать курсивом и маленькими буквами, чтобы их можно было отличить от кодируемых ими белков. Например, белок FOG-2 кодируется геном fog-2 .
. У гермафродитов белок HER-1 не образуется, но они все равно производят спермии, потому что активность гена tra-2 на стадии личинки подавляется белками FOG-2 и GLD-1. Таким образом, гермафродиты получаются из-за введения в игру дополнительного правила, связанного с регуляцией гена tra-2 .
Запутались? Ничего страшного, это в порядке вещей. Регуляторные сети, управляющие развитием животных, как правило, сложны, громоздки и неоптимальны [30] Почему естественный отбор делает их именно такими — разговор отдельный; в книге «Рождение сложности» мы уже начали обсуждать эту тему; мы вернемся к ней в главе 8, а здесь лишь напомним, что в природе всегда есть место совершенствованию, это не идеальный священный текст.
. К счастью, их не нужно учить наизусть никому, кроме студентов соответствующих специальностей. Чтобы облегчить дело, можно представить устрашающее описание из предыдущего абзаца в виде графической схемы (см. следующую страницу).
На таких схемах стрелочками обозначают положительные влияния (включение гена, активация белка), а «тупичками» — отрицательные (ингибирование белка, подавление активности гена).
Так вот, зная все это, исследователи подумали, что можно попробовать превратить самок раздельнополого вида C. remanei в гермафродитов, подавив активность какого-нибудь гена, блокирующего сперматогенез. Например, гена tra-2 .
Начали они с того, что отключили tra-2 при помощи РНК-интерференции [31] РНК-интерференция — метод отключения генов, основанный на использовании одного из защитных механизмов клетки. Вводят в клетку двухцепочечную РНК с такой же последовательностью нуклеотидов, как у целевого гена. Клетка «думает», что в нее проник вирус, и начинает уничтожать все РНК с такой последовательностью нуклеотидов — в том числе матричные РНК, считанные с интересующего нас гена.
. В результате из яиц с двумя X-хромосомами развились вместо самок обыкновенные самцы, производящие нормальные спермии, но никаких яйцеклеток.
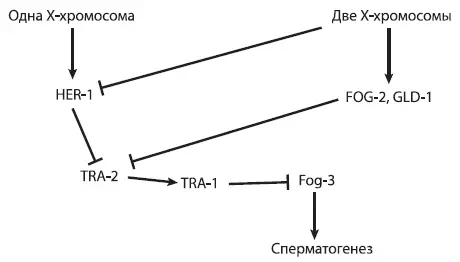
Схема регуляции развития пола у нематоды C. elegans , у которой вместо самок гермафродиты.
Тогда ученые справедливо рассудили, что они, видимо, перестарались. Ведь у настоящих гермафродитов активность гена tra-2 хоть и снижена, но не до нуля. Когда эту ситуацию удалось воспроизвести у C. remanei , уменьшив экспрессию tra-2 , на свет появились животные, которых авторы назвали псевдогермафродитами . Тело у них было «женское», но в гонадах формировались одновременно и яйцеклетки, и спермии. Правда, эти существа оказались неспособны к самооплодотворению (отсюда и приставка «псевдо»). После скрещивания с самцом превдогермафродиты откладывали оплодотворенные яйца, хоть и в меньшем количестве, чем обычные самки. Это значит, что яйцеклетки у них более или менее нормальные, а неспособность к самооплодотворению, скорее всего, объясняется дефектами спермиев.
Действительно, спермии псевдогермафродитов оказались нормальны по всем параметрам, кроме одного: они неактивны, никуда не ползут, и в частности не перебираются в сперматеки.
У обычных нематод спермии активируются (начинают ползать) под воздействием белков, содержащихся в семенной жидкости. Сохранили ли спермии псевдогермафродитов способность активироваться под действием этих белков? Чтобы это проверить, авторы скрещивали псевдогермафродитов с самцами C. elegans . Самцы C. elegans охотно спариваются с самками C. remanei , но эти браки бесплодны (как-никак два вида разошлись, судя по показаниям молекулярных часов, около 80 млн лет назад, а для развития генетической несовместимости обычно хватает нескольких миллионов лет [32] О механизмах развития несовместимости мы поговорим в главе 6, посвященной видообразованию.
). Идея такого спаривания как раз и состояла в том, чтобы половые клетки от разных видов не сливались, но при этом псевдогермафродиты могли получить нормальные белки семенной жидкости.
Интервал:
Закладка:








