Александр Марков - Эволюция. Классические идеи в свете новых открытий

- Название:Эволюция. Классические идеи в свете новых открытий
- Автор:
- Жанр:
- Издательство:АСТ: CORPUS
- Год:2014
- Город:Москва
- ISBN:978-5-17-083218-7
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Александр Марков - Эволюция. Классические идеи в свете новых открытий краткое содержание
Книга Александра Маркова и Елены Наймарк рассказывает о новейших исследованиях молекулярных генетиков и находках палеонтологов, которые дают ответы на эти и многие другие вопросы о видоизменениях в природе. Тысячи открытий, совершенных со времен Дарвина, подтверждают догадки родоначальников теории эволюции; новые данные ничуть не разрушают основы эволюционной теории, а напротив, лишь укрепляют их.
Александр Марков, заведующий кафедрой биологической эволюции биофака МГУ, и Елена Наймарк, ведущий научный сотрудник Палеонтологического института им. А. А. Борисяка, — известные ученые и популяризаторы науки. Двухтомник «Эволюция человека» (2011), написанный ими в соавторстве, стал настольной книгой не только для студентов и ученых-биологов, но и для множества людей за пределами профессионального сообщества.
Эволюция. Классические идеи в свете новых открытий - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
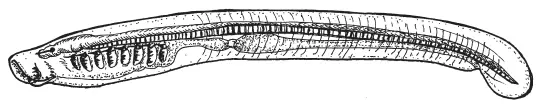
Пескоройка — личинка миноги — представляет собой, как и ланцетник, живую упрощенную схему типичного хордового. Она не сосет кровь у рыб, как многие взрослые миноги, а фильтрует воду через жаберные щели, подобно ланцетнику.
Как и следовало ожидать, каждая из 17 предковых хромосом превратилась в геноме его потомков (в том числе человека) в набор обрывков , рассеянных по разным хромосомам. Теперь мы подошли к самому главному. Оказалось, что каждый из 17 наборов в свою очередь подразделяется на четыре части. Например, набор, соответствующий предковой хромосоме № 8, в геноме человека состоит из девяти фрагментов ДНК. Они подразделяются на четыре группы (в трех группах по два фрагмента, в четвертой — три). Если сравнить между собой этих четыре группы фрагментов, то выясняется, что многие гены представлены в нескольких из них в виде паралогов(или паралогичных генов), по одному паралогу в группе. Паралогичные гены — это родственные гены, когда-то произошедшие от общего гена-предка в результате дупликации. Таким образом, каждая предковая хромосома превратилась в геноме позвоночных в четыре сходные между собой группы фрагментов ДНК . Сходство заключается в том, что если в одной группе есть какой-то ген, то в трех других с высокой вероятностью найдутся его паралоги. Это можно объяснить только тем, что на ранних этапах эволюции позвоночных действительно произошло учетверение генома — две последовательные дупликации. Гипотезу Оно, высказанную более 40 лет назад, наконец можно считать доказанной.
Дупликации произошли после того, как предки хордовых отделились от предков ланцетников и оболочников (их геномы не учетверены и не удвоены), и до того, как разошлись пути костных и хрящевых рыб. Скорее всего, первое удвоение произошло незадолго до, а второе сразу после разделения линий бесчелюстных (миноги, миксины) и челюстноротых (рыбы и наземные позвоночные). Эти линии разделились в раннем кембрии, примерно 530 млн лет назад.
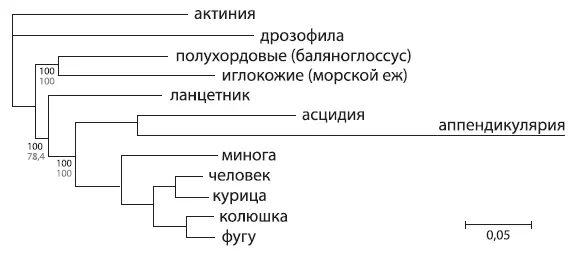
Эволюционное дерево вторичноротых, основанное на последовательностях 1090 белков. Актиния и дрозофила использованы в качестве «внешних групп», асцидия и аппендикулярия представляют оболочников. Длина ветвей отражает количество аминокислотных замен. Из Putnam et al., 2008 .
Удвоение целого генома — далеко не уникальный случай в истории жизни. Еще одна полногеномная дупликация произошла 350 млн лет назад у предков костистых рыб; 40 млн лет назад то же самое случилось в одной из групп лягушек. Обычно после этого большинство «лишних» генов быстро теряется, и лишь некоторым из них находится новое применение. До тех пор пока два гена-паралога не начнут выполнять в организме разные функции, естественный отбор не может воспрепятствовать мутационной поломке или утрате одного из них.
Ученые составили список из 8437 генов, которые были у общего предка хордовых и сохранились хотя бы у некоторых современных позвоночных. Все эти гены в свое время учетверились, но большая часть появившихся паралогов была утрачена. Сохранились паралоги у 20–25 % генов. Среди генов, дубликаты которых «пригодились» ранним позвоночным, резко повышена доля тех, которые участвуют в регуляции транскрипции (гены — регуляторы активности других генов), в работе нервной системы, в регуляции эмбрионального развития и в передаче разнообразных сигналов. Это явно неслучайный выбор. Он хорошо согласуется с идеей о том, что геномные дупликации у предков позвоночных стали предпосылкой для последующей эволюции по пути усложнения.
Но повторим — естественный отбор не умеет заглядывать в будущее. Он не мог заранее знать, что позвоночные собираются эволюционировать именно в сторону усложнения. Здесь уместно вспомнить о механизме «ухода от адаптивного конфликта», о котором мы говорили выше. Не потому ли сохранились дубликаты генов-регуляторов, что уже у древнейших позвоночных эти гены были многофункциональными и не могли толком оптимизироваться для выполнения какой-то одной формообразующей функции? По-видимому, полногеномные дупликации предоставили позвоночным уникальную возможность быстро снять многие из накопившихся адаптивных конфликтов и, как следствие, избавиться от множества запретов, стоявших на пути морфологических преобразований.
Откуда взялся новый фермент?
Амплификация (увеличение числа копий гена; частным случаем амплификации является дупликация — удвоение) с последующим разделением функций между копиями — по-видимому, самый распространенный способ появления новых генов. Мы уже знаем, что новая функция может появиться у одной из копий как до, так и после амплификации. В обоих случаях обязательным этапом является закрепление полезных мутаций в одной или нескольких копиях размножившегося гена. Проблема в том, что полезные мутации должны появиться быстро — до того как избыточные копии гена будут безнадежно испорчены вредными мутациями. Иначе все вернется на круги своя и в геноме снова останется только одна рабочая копия гена плюс некоторое количество «мусора»— поврежденных мутациями псевдогенов(вышедших из строя, неработающих генов), в которые превратятся остальные копии.
Биологи из Швеции и США разработали теоретически ( Bergthorsson et al., 2007 ), а затем и экспериментально подтвердили ( Näsvall et al., 2012 ) сценарий появления новых генов, который они назвали ИАД (инновация — амплификация — дивергенция). Сценарий похож на уже знакомый нам уход от адаптивного конфликта, но имеет важное отличие. В сценарии ИАД амплификация сама по себе имеет адаптивный смысл: новообразованные копии гена с самого начала не являются избыточными. Они полезны, и поэтому отбор препятствует их порче. Это дает им время дождаться появления полезных мутаций. Главная хитрость тут в том, что амплификация гена увеличивает производство кодируемого им белка, а это, как правило, усиливает выполнение функций, за которые отвечает белок.
Схема ИАД показана на рисунке. У гена с основной функцией А появляется (или давно существует в качестве «побочного эффекта») дополнительная функция B, которая поначалу осуществляется с низкой эффективностью (и потому обозначается как b). Если эта побочная функция вдруг окажется полезной (например, из-за изменения условий среды), то отбор начнет поддерживать мутации, усиливающие эту функцию. Простейшим способом добиться этой цели, не нарушив функцию А, является амплификация бифункционального гена. Чем больше копий гена будет в геноме, тем больше будет молекул соответствующего белка и тем эффективнее станет осуществляться функция B. Таким образом, отбор будет поддерживать дупликации гена и защищать появляющиеся копии от мутационных повреждений. Заметим, что амплификация генов — весьма распространенная категория мутаций. Например, у бактерии Salmonella enterica , с которой работали исследователи, вероятность дупликации любого гена составляет примерно 10 −5на каждое клеточное деление [72].
Читать дальшеИнтервал:
Закладка:








