Александр Марков - Эволюция. Классические идеи в свете новых открытий

- Название:Эволюция. Классические идеи в свете новых открытий
- Автор:
- Жанр:
- Издательство:АСТ: CORPUS
- Год:2014
- Город:Москва
- ISBN:978-5-17-083218-7
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Александр Марков - Эволюция. Классические идеи в свете новых открытий краткое содержание
Книга Александра Маркова и Елены Наймарк рассказывает о новейших исследованиях молекулярных генетиков и находках палеонтологов, которые дают ответы на эти и многие другие вопросы о видоизменениях в природе. Тысячи открытий, совершенных со времен Дарвина, подтверждают догадки родоначальников теории эволюции; новые данные ничуть не разрушают основы эволюционной теории, а напротив, лишь укрепляют их.
Александр Марков, заведующий кафедрой биологической эволюции биофака МГУ, и Елена Наймарк, ведущий научный сотрудник Палеонтологического института им. А. А. Борисяка, — известные ученые и популяризаторы науки. Двухтомник «Эволюция человека» (2011), написанный ими в соавторстве, стал настольной книгой не только для студентов и ученых-биологов, но и для множества людей за пределами профессионального сообщества.
Эволюция. Классические идеи в свете новых открытий - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Таким образом, модель Добжанского — Мёллера предсказывает, что число генов, ответственных за генетическую несовместимость, должно расти с ускорением, как снежный ком. Если предположить, что различия накапливаются с постоянной скоростью (т. е. исходить из допущения о равномерном ходе молекулярных часов), то число «генов несовместимости» должно быть пропорционально квадрату времени, прошедшего с момента разделения популяций.
Модель предполагает, что главную роль в формировании постзиготической изоляции играют попарные антагонистические взаимодействия («антагонистический эпистаз») между разными локусами. Если учитывать возможность конфликтов не только между двумя, но и между тремя и более локусами, то картина существенно усложнится.
В течение 70 с лишним лет, прошедших с момента появления этой модели, в ее работоспособности мало кто сомневался. Ее преподавали в университетах, а генетики, опираясь на нее, занимались поиском конкретных «генов несовместимости», порой весьма успешно. Но вот проверить ее напрямую не удавалось. Для этого нужно сравнить число генетических различий с числом «генов несовместимости» в нескольких парах родственных видов, различающихся по времени своего расхождения. Главная проблема в том, как подсчитать «гены несовместимости».
Лишь в 2010 году сразу двум научным коллективам одновременно удалось преодолеть эту трудность. Первая работа выполнена сотрудниками Чикагского университета на дрозофилах ( Matute et al., 2010 ). Среди ее авторов — Джерри Койн, написавший замечательную научно-популярную книгу Why evolution is true ( Coyne, 2009 ) о доказательствах эволюции.
Ученые воспользовались тем обстоятельством, что некоторые виды дрозофил, хотя и разошлись давно, все еще сохранили способность к гибридизации. Авторы скрещивали самок Drosophila melanogaster с самцами двух других видов: D. simulans и D. santomea . Эволюционные пути видов melanogaster и simulans разошлись 5,4 млн лет назад, melanogaster и santomea — примерно 12,8 млн лет назад. Таким образом, время независимого существования в двух парах видов различается в 2,4 раза, и количество генетических различий тоже (собственно, по последнему и вычислили первое!). Если гипотеза Добжанского — Мёллера верна, число «генов несовместимости» в паре melanogaster — santomea должно быть примерно в 2,4 2 = 5,76 раза больше, чем в паре melanogaster — simulans .
Чтобы подсчитать «гены несовместимости» в каждой паре, авторы использовали большую, включающую сотни линий, коллекцию дрозофил D. melanogaster с различными делециями — выпадениями небольших участков генома. У мух из этих линий делеции поддерживаются в гетерозиготном состоянии. То есть у каждой мухи в одной хромосоме отсутствует какой-то кусочек, но в другой, парной хромосоме этот кусочек имеется. При этом вторая, целая хромосома, во-первых, «помечена» какой-нибудь доминантной мутацией (чтобы сразу было видно, кому из потомства она досталась), во-вторых, она неспособна рекомбинировать — обмениваться участками с парной хромосомой во время мейоза. Половина половых клеток такой мухи получает целую хромосому с меткой, а другая половина — хромосому с недостающим фрагментом и без метки.
Делеции, имеющихся в разных линиях, в общей сложности покрывают почти весь геном. Авторы скрещивали самок из этих линий (напомним, все они относятся к виду melanogaster ) с самцами simulans и santomea . От таких межвидовых скрещиваний рождаются только самки. Потомство мужского пола погибает на ранних стадиях.
Здесь нужно сделать небольшое отступление, чтобы объяснить, почему так получается. Дело в том, что у самцов дрозофил, как и у людей, только одна X-хромосома, а у самок — две. Поэтому гены, расположенные в X-хромосоме, у самцов имеются в единственном экземпляре. Если хоть в одном из них имеется летальная мутация, самец обречен. У самки же есть вторая копия X-хромосомы, в которой данный ген может быть нормальным. Нормальная копия гена «перекрывает» действие испорченной копии, и муха выживает.
Что же за летальные мутации сидят в генах Х-хромосомы гибридных самцов? Это, очевидно, те самые «гены несовместимости», ради поиска которых все и было затеяно. Они конфликтуют с какими-то другими генами в геноме того вида, которому не принадлежит данная X-хромосома. Причем из того факта, что гибридные самки все-таки выживают, можно сделать вывод, что все «гены несовместимости» в данной паре видов рецессивны. Допустим, если несовместимыми являются гены A 2и B 2, причем ген B расположен на Х-хромосоме, то гибридные самки с генотипом A 1A 2B 1B 2будут жить, а самцы с генотипом A 1A 2B 2погибнут.
Итак, при скрещивании самки melanogaster с самцом simulans или santomea все потомство состоит из дочерей. Причем эти дочери будут двух типов. Первому типу достанется от матери, самки melanogaster , хромосома с отсутствующим кусочком. У таких гибридных мух соответствующий кусочек будет присутствовать в единственном экземпляре — в парной хромосоме, полученной от отца. Таким образом, отцовские гены, расположенные в этом кусочке, окажутся в «оголенном» ( гемизиготном) состоянии — как если бы они сидели в X-хромосоме самца. Если в этом фрагменте хромосомы имеется рецессивный «ген несовместимости», гибридные самки будут иметь пониженную жизнеспособность или вообще погибнут. Второй тип дочерей получит от матери melanogaster целую хромосому с меткой (по метке можно определить, к какому из двух типов относится данная гибридная самка). Все гены будут в двух экземплярах, и рецессивная несовместимость не проявится, т. е. все самки второго типа должны выжить.
Таким образом, по количественному соотношению гибридных дочерей первого и второго типа можно определить, содержится ли в данном кусочке хромосомы «ген несовместимости». Если обоих типов примерно поровну — значит, «гена несовместимости» там нет; если самок первого типа оказалось существенно меньше, чем второго, значит, он там есть.
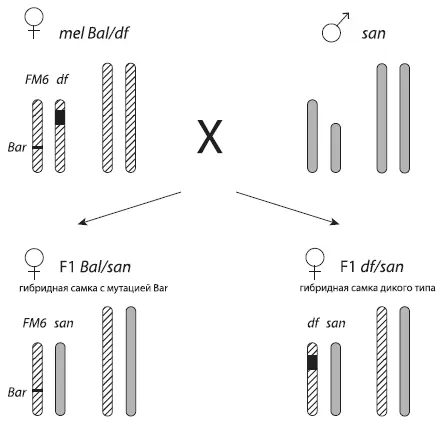
Схема экспериментов с дрозофилами. Хромосомы melanogaster (mel) показаны косой штриховкой, хромосомы santomea (san) — серым цветом. Скрещивается самка melanogaster с самцом santomea. Самая короткая — Y -хромосома , X -хромосома подлиннее. Черным показан отсутствующий участок хромосомы (в данном случае — X -хромосомы). Хромосома с отсутствующим участком помечена буквами df (от deficiency ). Вторая X -хромосома самки melanogaster — целая и помеченная доминантной мутацией Bar . В результате межвидового скрещивания получаются гибридные дочери двух типов: одни несут «целую» материнскую хромосому с меткой, другие получают от матери «урезанную» хромосому df . По выживаемости дочерей второго типа можно понять, есть ли в том кусочке ДНК, который был вырезан из хромосомы df , «ген несовместимости». Из Matute et al., 2010 .
Читать дальшеИнтервал:
Закладка:








