Александр Марков - Эволюция. Классические идеи в свете новых открытий

- Название:Эволюция. Классические идеи в свете новых открытий
- Автор:
- Жанр:
- Издательство:АСТ: CORPUS
- Год:2014
- Город:Москва
- ISBN:978-5-17-083218-7
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Александр Марков - Эволюция. Классические идеи в свете новых открытий краткое содержание
Книга Александра Маркова и Елены Наймарк рассказывает о новейших исследованиях молекулярных генетиков и находках палеонтологов, которые дают ответы на эти и многие другие вопросы о видоизменениях в природе. Тысячи открытий, совершенных со времен Дарвина, подтверждают догадки родоначальников теории эволюции; новые данные ничуть не разрушают основы эволюционной теории, а напротив, лишь укрепляют их.
Александр Марков, заведующий кафедрой биологической эволюции биофака МГУ, и Елена Наймарк, ведущий научный сотрудник Палеонтологического института им. А. А. Борисяка, — известные ученые и популяризаторы науки. Двухтомник «Эволюция человека» (2011), написанный ими в соавторстве, стал настольной книгой не только для студентов и ученых-биологов, но и для множества людей за пределами профессионального сообщества.
Эволюция. Классические идеи в свете новых открытий - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Допустим, у нас есть маленькая популяция мюмзиков из 40 особей. Для простоты примем, что мюмзики гаплоидны, т. е. имеют одинарный набор хромосом — один-единственнный экземпляр генома, а не два, как у нас с вами, диплоидныхорганизмов. Достигнув возраста в один год, каждый мюмзик рожает нескольких детенышей, а потом сразу умирает от старости. Все детеныши одинаковые, но выжить из них могут только 40 — больше мюмзиков просто не помещается в старой железной бочке на дне пруда, где живет изучаемая популяция. Смертность детенышей случайна — ровно 40 случайно выбранных везунчиков станут взрослыми и через год произведут потомство.
Поскольку смертность случайна, приспособленность мюмзиков определяется только их плодовитостью, т. е. числом детенышей. Плодовитость зависит от генотипа. Допустим, у мюмзика есть один ген, влияющий на плодовитость. Обозначим его буквой А. Между прочим, не смейтесь: такая модель вполне годится для изучения некоторых законов популяционной генетики.
Некогда у всех мюмзиков был только один вариант (аллель) гена А. Обозначим его А 1. Но потом у одной особи произошла мутация, и в результате появился второй вариант гена — аллель А 2. Допустим (опять-таки для простоты), что в нашей популяции, когда мы приступили к ее изучению, у половины мюмзиков ген А был представлен первым вариантом, у другой половины — вторым. Таким образом, частота аллеля А 2равна 0,5 (q 2 = 0,5) и частота аллеля А 1такая же (q 1 = 0,5).
Нам нужно ответить на вопрос: как будет меняться со временем частота аллеля A 2, если мутация, которая привела к его возникновению, была нейтральной?
Раз мутация была нейтральной, значит, плодовитость обладателей обоих аллелей одинакова. Допустим, они все рожают ровно по десять детенышей. Разумеется, потомство наследует родительский аллель гена А.
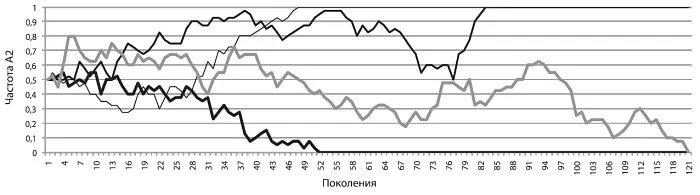
Может показаться, что раз аллели имеют одинаковую приспособленность, то их так и останется поровну. Это неверный ответ. На рисунке [см. ниже] показан результат четырех запусков нашей модели. Мы видим, что частота А 2во всех четырех случаях хаотически колебалась (по-научному такие колебания как раз и называются случайными блужданиями). Блуждания продолжаются до тех пор, пока частота аллеля не упрется либо в верхнюю «точку невозврата» (q 2 = 1, частота аллеля достигла 100 %, аллель зафиксировалсяв генофонде), либо в нижнюю (q 2 = 0, частота упала до нуля, аллель элиминировалсяиз генофонда).
Так бывает всегда. Если аллель нейтрален, его частота будет «случайно блуждать» между нулем и единицей до тех пор, пока не упрется либо в верхний, либо в нижний предел. Рано или поздно нейтральный аллель либо зафиксируется (достигнет частоты 1), либо элиминируется — исчезнет из генофонда. Третьего не дано. Таким образом, блуждания хоть и случайны, их исход предсказуем. Мы точно знаем, что дело кончится либо фиксацией, либо элиминацией. Чем больше популяция, тем в среднем дольше придется ждать исхода [17] Не вдаваясь в сложные вычисления, заметим лишь, что число поколений, которое (в среднем) должно пройти до фиксации или элиминации нейтральной мутации, имеет примерно тот же порядок величин, что и численность популяции. Если в популяции несколько тысяч особей, то и ждать придется несколько тысяч поколений, в миллионной популяции речь будет идти о миллионах поколений. Поэтому на судьбу нейтральных мутаций сильное влияние оказывают «бутылочные горлышки» — периоды сокращения численности популяции, во время которых нейтральный аллель может потеряться или зафиксироваться особенно быстро.
, но он все равно неизбежен.
Здесь самое время сказать, что случайные, не зависящие от приспособленности изменения частот аллелей называются генетическим дрейфом. Под властью дрейфа находятся все нейтральные мутации (а в маленьких популяциях отчасти также и вредные с полезными, но об этом позже).
Можно ли вычислить вероятность того, что нейтральный аллель в конце концов зафиксируется, а не элиминируется? Да, это просто. В нашем примере аллели А 1и А 2исходно находились в равном положении, потому что у обоих была частота 0,5. Очевидно, что в такой ситуации шансы на фиксацию у них должны быть одинаковыми и равными 0,5. В половине случаев А 1зафиксируется, А 2элиминируется, в половине — наоборот.
Ну а если конкурирующих нейтральных аллелей не два, а, скажем, четыре и начальные частоты у них тоже одинаковые (0,25)? В этом случае дело кончится фиксацией одного из аллелей и элиминацией трех остальных, причем для каждого аллеля вероятность фиксации равна 0,25. Таким образом, очевидно, что вероятность фиксации нейтральной мутации в будущем просто-напросто равна ее частоте в данный момент: P fix = q.
Если вы, дорогие читатели, еще не устали от этой примитивной математики, то позвольте познакомить вас еще с двумя простыми, интересными и полезными формулами.
Сколько нейтральных мутаций будет фиксироваться в популяции в каждом поколении? (Имеются в виду мутации уже не в одном и том же, а в разных генах.) Если мы сумеем это вычислить, то получим прекраснейший инструмент — молекулярные часы. Тогда мы сможем по количеству нейтральных генетических различий определить, когда жил последний общий предок сравниваемых организмов.
Вывод этой формулы — подлинный шедевр «биологической математики». Судите сами. Определим сначала, сколько новых мутаций появляется в популяции в каждом поколении. Будем считать для простоты, что подавляющее большинство мутаций нейтральны (это недалеко от истины). Ответ очевиден: U × N, где U — темп мутагенеза (среднее число новых мутаций у каждой новорожденной особи), N — численность популяции. Определить U можно, просто сравнивая геномы детей и родителей.
Теперь нужно понять, какая часть из этих U × N только что появившихся мутаций в итоге зафиксируется. Это и будет искомая величина — темп фиксации нейтральных мутаций в генофонде популяции за одно поколение. Обозначим ее буквой V.
Мы уже знаем, что вероятность фиксации мутации равна ее частоте: P fix = q. Чему же равна частота только что появившейся мутации? Это совсем просто. Поскольку мутация только что появилась, она пока есть только у одной особи. Следовательно, ее частота равна 1/N. Вот, собственно, и все. Умножаем количество новых мутаций на вероятность фиксации каждой из них (т. е. на долю мутаций, которые в итоге зафиксируются) и получаем ответ: V = U × N × 1/N. Поразительно! Численность популяции волшебным образом сокращается, и величина N уходит из уравнения. Мы приходим к выводу, что темп фиксации нейтральных мутаций не зависит от численности и равен просто-напросто скорости мутагенеза: V = U. Не знаю, как вы, дорогие читатели, а мы, биологи, приходим в восторг от такой красоты.
Читать дальшеИнтервал:
Закладка:








