Сергей Савельев - Происхождение мозга

- Название:Происхождение мозга
- Автор:
- Жанр:
- Издательство:Веди
- Год:2005
- ISBN:5-94624-025-0
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Сергей Савельев - Происхождение мозга краткое содержание
Происхождение мозга - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Следовательно, основной причиной появления нервных клеток было стремление «голодных» клеток покровов увеличить свой метаболизм. Следствием роста отростков стала интеграция внешней и внутренней информации организма об эффективности питания. В этой информации были заинтересованы все клетки этого древнего животного, что заставило их сформировать встречные отростки, направленные к пронейронам. Объединение пронейронов в сети дало огромный выигрыш в получении пищи и увеличении метаболизма животного. Организм смог не пассивно фильтровать воду, а активно находить богатые пищевые участки или поедать себе подобных. Началась конкурентная эволюция эффективности нервных систем, которая быстро превратилась в физическую битву морфологических конструкций. В кратчайшие по историческим меркам сроки были опробованы все мыслимые и немыслимые типы интегрирующих нервных систем. Они сталкивались между собой и порождали ещё более оригинальные и эффективные устройства. О них мы можем судить только по фантастическим вариантам строения нервных систем беспозвоночных, дошедших до наших времён.
Беспозвоночные были первыми многоклеточными обладателями нервной системы. Эволюция этой группы животных наполнена запутанными и драматическими коллизиями, которые пока не входят в круг нейробиологических интересов. Стараясь не вмешиваться в проблемы эволюции беспозвоночных, мы рассмотрим только самые общие принципы организации их нервной системы.
§ 20. Нервная система с радиальной симметрией
Наиболее простой вариант строения нервной системы мы встречаем у стрекающих (кишечнополостных). Как уже говорилось выше, их нервная система построена по диффузному типу. Клетки образуют пространственную сеть, которая расположена в мезоглее (рис. II-4, а). Небольшое скопление нервных клеток в окологлоточной области образует подобие распределённого нервного центра. Он интегрирует все несложные реакции тела кишечнополостных и является эволюционным предшественником ганглиозной нервной системы. В окологлоточном нервном кольце формируются параллельные ганглии, описанные в главе (см. рис. I-16). По-видимому, этот тип строения нервной сети был исходным для всех групп животных.
При всей кажущейся простоте диффузный тип нервной системы обеспечивает довольно сложное поведение кишечнополостных. Хорошо известно, что раки-отшельники используют актиний для защиты от хищников. Они выбирают наиболее подходящих особей и пересаживают их себе на раковину. Классическим примером является симбиоз актиний и рака-отшельника. Однако мало известно, что сами актинии также могут выбирать наиболее подходящую поверхность раковины и перемещаться на неё. Иначе говоря, актинии такие же активные, хотя и бессознательные, участники симбиоза, как и раки-отшельники (Холодковский, 1914; Meglitsch, Schram, 1991).
В скромных рамках диффузной нервной системы известно необычайно большое количество вариантов строения. Их всех объединяют радиальная или изоморфная симметрия и общая тенденция к объединению нервных клеток в некие скопления. С момента появления пронейральной сети у губкоподобных организмов началась дифференциальная концентрация нервных элементов. В начале эволюции многоклеточных животных появилось бесконечное разнообразие вариантов строения нервной системы, которые реализовались у кишечнополостных и частично сохранились до нашего времени (см. рис. II-4).
Нервные клетки концентрировались различными способами. Самым простым способом интеграции нервных сетей стало окологлоточное нервное кольцо. Его появление вполне оправдано тем, что оно находилось на границе поступления пищи в организм кишечнополостных. Пища была тем ведущим стимулом, который определял и оценивал успех морфологических изменений нервной системы. Тот, кто мог эффективнее контролировать поступление пищи, увеличивал свой метаболизм и репродуктивные возможности. Самым простым движением, позволяющим проиллюстрировать действие диффузной нервной системы, является реакция на механическое раздражение. Пресноводная гидра (Pelmatohydra oligactis) при малейшем раздражении сжимается в микроскопический комочек. Это происходит за счёт расположенных продольно в эктодерме и поперечно в энтодерме сократимых белков. Кроме генерализованной реакции, кишечнополостные могут дифференциально пользоваться отдельными щупальцами или их группами. Гидры способны передвигаться, чередуя при переворотах опору на подошву и ротовое отверстие.
Тем не менее диффузная сеть с окологлоточным нервным кольцом была относительно медленно действующим устройством. Измеренная проводимость по нервной сети кишечнополостных составляет не более 5-20 см/с. Этого явно не хватает животным размером более 5 см, поэтому уже у актиний выделились участки нервной сети с высокой скоростью проведения (см. рис. II-4, в). В некоторых случаях она достигает
см/с, что делает актиний изощрёнными охотниками за значительно более эволюционно продвинутыми позвоночными. Окологлоточное нервное кольцо было явным достижением, но оно не могло дифференциально управлять всем телом или обеспечить контроль за свободным плаванием.
Среди предков современных одиночных актиний явно были свободноплавающие существа. На это указывает двойная нервная сеть в их теле (см. рис. II-4, б). Одна диффузная сеть расположена под эктодермой в мезоглее и ничем не отличается от таковой у других кишечнополостных (см. рис. II-4, а). Другая нервная сеть лежит в той же мезоглее, но уже около энтодермы. Они связаны между собой только в зоне окологлоточного нервного кольца, которое начинает играть как интегрирующую, так и разделяющую роль. По-видимому, такие двойные сети возникли на заре эволюции нервной системы и были нужны для активного свободного плавания. Животное с автономной «эктодермальной» сетью могло активно двигаться в толще воды. Сокращение эктодермальных клеток позволяло животному перемещаться, не вовлекая в этот процесс пищеварительную систему.
Рис. II-4. Предполагаемые первые этапы (показано стрелками) усложнения строения нервной системы кишечнополостных с радиальной симметрией.
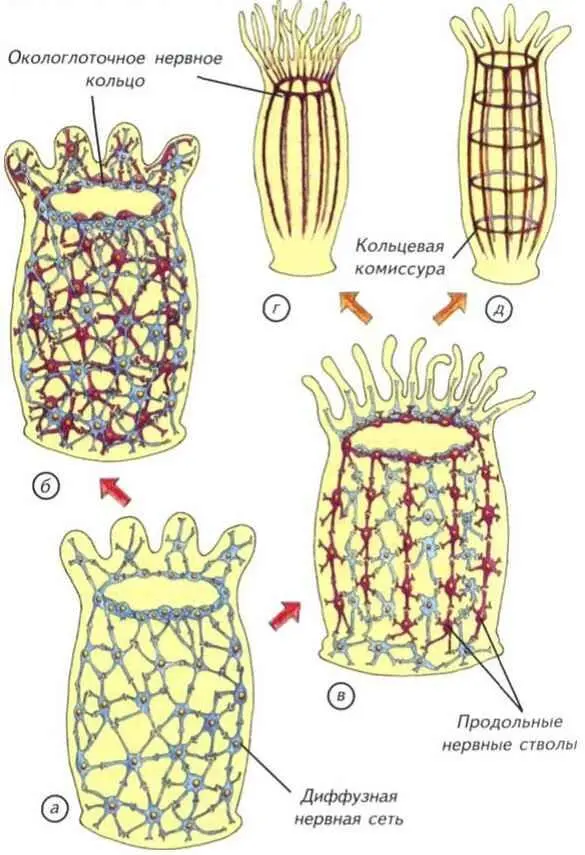
а — однослойная сеть примитивных гидроидов; б — двойная нервная сеть актиний; в — нервная сеть актиний со скоростными проводящими цепями клеток; г — нервная сеть восьмилучевого полипа; д — нервный аппарат радиально-комиссурального типа.
Не исключено, что мезоглея была у этих существ намного толще и рыхлее. Пищеварительная нервная сеть с энтодермальными сократимыми клетками функционировала относительно автономно, перемещая пищевые частицы без эктодермальных сокращений. Такой самодвижущийся пылесос мог быть крайне эффективным при избытке мелких пищевых частиц. Эволюционные преимущества подвижных фильтраторов хорошо известны, поскольку усатые киты являются самыми крупными животными на планете.
Читать дальшеИнтервал:
Закладка:



