Дэвид Хьюбел - Глаз, мозг, зрение

- Название:Глаз, мозг, зрение
- Автор:
- Жанр:
- Издательство:Мир
- Год:1990
- Город:Москва
- ISBN:5-03-001254-0
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Дэвид Хьюбел - Глаз, мозг, зрение краткое содержание
В книге известного американского нейрофизиолога, лауреата Нобелевской премии, обобщены современные представления о том, как устроены нейронные структуры зрительной системы, включая кору головного мозга, и как они перерабатывают зрительную информацию. При высоком научном уровне изложения книга написана простым, ясным языком, прекрасно иллюстрирована. Она может служить учебным пособием по физиологии зрения и зрительного восприятия.
Для студентов биологических и медицинских вузов, нейрофизиологов, офтальмологов, психологов, специалистов по вычислительной технике и искусственному интеллекту.
Глаз, мозг, зрение - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
В отличие от клеток на более низких уровнях зрительной системы нейроны, избирательно чувствительные к ориентации стимула, гораздо лучше отвечают на движущиеся, чем на неподвижные линии. Именно поэтому (см. рис. 41) при стимуляции таких нейронов мы использовали линии, движущиеся через рецептивное поле. Если использовать в качестве стимула неподвижную мелькающую (периодически вспыхивающую) линию, то зачастую клетка дает слабый ответ, и в этом случае предпочтительной оказывается такая же ориентация, как и при движущейся линии.
Многие клетки (вероятно, треть всей популяции) дают еще один характерный вид ответа на движущийся стимул. Вместо того чтобы давать один и тот же импульсный разряд независимо от направления движения, такие клетки отвечают более энергично при одном определенном направлении. Бывает даже так, что движение в одну сторону вызывает сильно выраженный ответ, а при движении в противоположную сторону нет вообще никакой реакции (это показано на рис. 41).
За один эксперимент можно оценить реакции 200–300 клеток, если после полного исследования одной клетки просто продвигать микроэлектрод дальше, до следующей клетки. Недостаток такой методики в том, что в одном прогоне можно исследовать только клетки, лежащие в коре на одной прямой линии: как только вы ввели тончайший микроэлектрод в корковую ткань, вы уже не можете сдвинуть его в поперечном направлении, не повредив при этом сам электрод или еще более нежную нервную ткань. С помощью такой методики регистрации самое большее, что мы можем сделать — это исследовать при одной проходке микроэлектрода примерно 50 клеток на миллиметр. Когда мы исследуем ориентационную избирательность нескольких сотен или тысяч клеток, оказывается, что все ориентации стимула встречаются примерно одинаково часто — вертикальная, горизонтальная и все промежуточные, наклонные ориентации. Если учесть характерные особенности окружающего нас мира, в который входят как деревья, так и линия горизонта, то возникнет вопрос: нет ли каких-нибудь выделенных ориентаций, таких как вертикаль и горизонталь, которые встречались бы чаще, чем другие? При попытках ответить на этот вопрос в разных лабораториях получали несколько разные результаты. Однако все исследователи соглашаются, что, если такие предпочтения действительно имеют место, они должны быть очень малы — настолько малы, что для выявления их требуется статистическая обработка данных. А в этом случае они вряд ли имеют какое-либо значение!
В стриарной коре обезьян примерно 70–80% клеток обладают свойствами ориентационной избирательности. Что касается кошек, то у них, по-видимому, все корковые клетки чувствительны к ориентации стимула, даже те, которые имеют прямые входы от наружных коленчатых тел.
Мы обнаружили заметные различия среди ориентационно-специфичных клеток, причем не столько в оптимальной ориентации стимула или в положении рецептивного поля на сетчатке, сколько в характере поведения клеток. Наиболее существенно различие между двумя классами нейронов — простыми и сложными клетками. Как можно догадаться по их названиям, клетки этих двух классов различаются по сложности своих ответных реакций. Поэтому мы сделали естественное предположение о том, что клетки с более простым поведением расположены в нейронной структуре коры ближе к ее входу.
В большинстве случаев по ответам простых клеток на стимул в виде маленького светового пятна можно предсказать их реакцию на стимул сложной формы. Каждая из простых клеток, подобно ганглиозным клеткам сетчатки, клеткам НКТ и корковым клеткам с центрально-симметричными рецептивными полями, имеет небольшое четко очерченное рецептивное поле. Предъявление в пределах этого рецептивного поля стимула в виде светового пятнышка вызывает либо on-, либо off-реакцию в зависимости от того, в какой именно участок рецептивного поля подан стимул. Различие между простыми клетками и клетками предыдущих уровней заключается в конфигурации зон возбуждения и торможения. На предыдущих уровнях это центрально-симметричная конфигурация — имеется одна центральная on- или off-зона (возбуждающая или тормозная) и окружающая ее со всех сторон кольцевая зона с противоположными свойствами (тормозная или возбуждающая). Простые клетки коры более сложны. Зоны возбуждения и торможения в их рецептивных полях всегда разделены одной прямой линией или двумя параллельными линиями (рис. 42). Чаще всего встречается такая конфигурация, когда к длинной и узкой возбуждающей зоне с двух сторон примыкают более широкие тормозные зоны (рис. 42, А ).
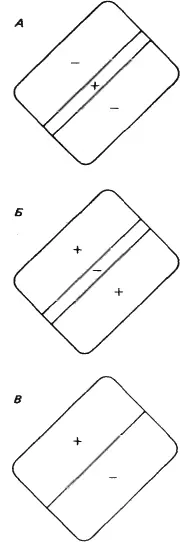
Рис. 42. Карты трех типичных рецептивных полей простых клеток. Оптимальными стимулами служили: для клетки А — светлая полоса против возбуждающей области ( +); для клетки Б — темная линия, покрывающая тормозную зону ( –); для клетки В — резкая граница «темное — светлое», совпадающая с границей между возбуждающей и тормозной зонами.
Для того чтобы проверить предполагаемую карту рецептивного поля, составленную путем тестирования маленьким световым пятном, мы попробовали использовать в качестве стимулов другие конфигурации. Вскоре мы выяснили, что чем большую долю той или иной зоны рецептивного поля покрывает данный стимул, тем сильнее выражено возбуждение клетки или ее торможение. Иными словами, имеет место пространственная суммация локальных воздействий. Мы обнаружили также явление антагонизма — взаимного погашения локальных воздействий при одновременной стимуляции возбуждающей и тормозной зоны. Таким образом, для клетки с рецептивным полем, изображенным на рис. 42, А, наиболее подходящим будет стимул в виде узкой полосы, расположенной в рецептивном поле так, чтобы она точно совпадала с возбуждающей зоной и не заходила в тормозную зону (см. рис. 43). Даже незначительное изменение ориентации этой полосы приведет к уменьшению эффективно действующей площади зоны возбуждения и затронет также тормозную зону; в результате частота разряда в ответе клетки уменьшится.
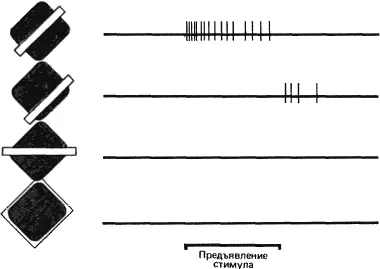
Рис. 43. Стимулы разных конфигураций вызывают различные реакции клетки с рецептивным полем такого типа, как А на рис. 42. Отрезком жирной линии внизу указан период (1 секунда), когда был включен стимул — светлая полоса. В первом случае (верхняя запись) показан ответ клетки на полосу оптимальных размеров, положения и ориентации. Во втором случае та же самая полоса покрывает только часть тормозной зоны (поскольку эта клетка не обладает спонтанной активностью, которая могла бы подавляться при торможении, здесь виден только разряд клетки при выключении стимула). В третьем случае полоса ориентирована так, что покрывает только малую часть возбуждающей зоны и соответственно малую часть тормозной зоны, и поэтому клетка не отвечает вообще. На нижней записи показан случай равномерного освещения всего рецептивного поля: ответа здесь тоже нет.
Читать дальшеИнтервал:
Закладка:







![Дэвид Доу - Почини свой мозг. Программа восстановления нейрофункций после инсульта и других серьезных заболеваний [litres]](/books/1143582/devid-dou-pochini-svoj-mozg-programma-vosstanovlen.webp)

