Александр Уголев - Естественные технологии биологических систем

- Название:Естественные технологии биологических систем
- Автор:
- Жанр:
- Издательство:Наука
- Год:1987
- Город:Ленинград
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Александр Уголев - Естественные технологии биологических систем краткое содержание
Книга посвящена концепции естественных технологий живых систем на различных уровнях организации последних и изложению доказательств, позволяющих преодолеть противопоставление естествознания и технологии. Эта концепция обосновывается на примере наиболее важных процессов в живых системах, их эволюции и происхождения. Охарактеризованы некоторые закономерности, которые могут быть интерпретированы как общие для естественных технологий живой природы и производственных технологий. Показано, что такие подходы плодотворны для понимания биологии в целом, процессов, протекающих в живых системах различной сложности, взаимодействий естественных и производственных технологий, в частности в медицине, экологии, питании и т.д.
Естественные технологии биологических систем - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Перечисленные экзогликозидазы синтезируются в кишечных клетках как хордовых, так и беспозвоночных животных. Важно, что внутриклеточные гликозидазы беспозвоночных животных, у которых имеет место внутриклеточное пищеварение, по своим функциям и многим характеристикам сходны с таковыми внеклеточных ферментов.
Что касается переваривания целлюлозы, составляющей большую часть пищи травоядных, то лишь небольшое число организмов обладает ферментными системами (КФ 3.2.1.4, целлюлаза), которые могут расщеплять это вещество на более простые продукты, способные к всасыванию и дальнейшему усвоению. В эту группу входит ряд брюхоногих и пластиножаберных моллюсков, некоторые ракообразные, рептилии и рыбы, насекомые, а также простейшие, использующие целлюлазу для деградации клеточных стенок зеленых водорослей и утилизации их содержимого. Целлюлаза играет важную роль в переваривании целлюлозы у кольчатых червей и иглокожих. Большинство травоядных животных (как позвоночных, так и беспозвоночных) для гидролиза целлюлозы используют симбионтные организмы. Кроме того, целлюлаза обнаружена у гнилостных бактерий и грибов.
Ферменты как внеклеточные, например панкреатическая липаза и кишечная моноглицеридлипаза хордовых, так и липазы беспозвоночных животных, реализующие внутриклеточное пищеварение, расщепляют жиры. Липазы гидролизуют преимущественно триглицериды с образованием 2-моноглицеридов и жирных кислот, при этом активирующее действие оказывают соли желчных кислот.
Панкреатическая фосфолипаза (КФ 3.1.1.4) секретируется в виде профермента профосфолипазы А и активируется трипсином. Фермент гидролизует эфирную связь глицерина и- жирной кислоты у второго атома углерода, превращая лецитин в изолецитин и жирную кислоту. Кишечная моноглицеридлипаза (КФ 3.1.1.23) гидролизует эфирные связи 2-моноглицеридов, образующихся при гидролизе триглицеридов. В гидролизе жиров у беспозвоночных животных, по-видимому, участвуют преимущественно эстеразы. Эти ферменты (кроме липазы) наблюдаются в средней кишке ракообразных, у насекомых, двустворчатых моллюсков, у которых встречаются как вне-, так и внутриклеточные пищеварительные эстеразы, у брюхоногих моллюсков, в кишечнике многощетинковых червей и иглокожих, а также у простейших.
Кроме указанных групп ферментов, существует несколько других, также играющих важную роль в пищеварительных процессах. Эти ферменты известны главным образом у хордовых животных, но, возможно, существуют также и у беспозвоночных. В качестве примера могут быть приведены щелочная фосфатаза (КФ 3.1.3.1), гидролизующая моноэфиры ортофосфор-ной кислоты, а также нуклеазы (РНКаза и ДНКаза), нуклеотидазы, нуклеозидазы и другие (КФ 3.1.3.—), продуцируемые клетками поджелудочной железы и синтезируемые клетками кишечного эпителия хордовых животных. Эти ферменты расщепляют полинуклеотиды и нуклеиновые кислоты пищи до пуринов, пиримидинов, дезоксирибозы, рибозы и фосфата.
Основные процессы гидролиза и транспорта у высокоорганизованных животных и человека, а также у ряда беспозвоночных реализуются в кишечнике. В связи с этим рассмотрим особенности структуры пищеварительных ферментов апикальной мембраны кишечных клеток млекопитающих, которые осуществляют заключительные этапы гидролиза основных групп пищевых веществ. При этом возможна прямая экстраполяция характеристик кишечных мембранных ферментов теплокровных организмов на характеристики ферментов более низкоорганизованных животных, так как строение мембраны и ферментативно активных белков, встроенных в фосфолипидный матрикс мембраны, практически универсально.
Большинство кишечных ферментов является трансмембранными интегральными белками (точнее, гликопротеинами) с большой молекулярной массой. Так, молекулярная масса кишечной щелочной фосфатазы — около 120 000—130 000 дальтон, аминопептидазы — варьирует от 225 000 до 280 000 в зависимости от вида животного, карбогидраз, в том числе сахаразно-изомальтазного комплекса, более 200 000. Однако некоторые ферменты, например дипептидаэы апикальной мембраны кишечных клеток, могут быть периферическими интегральными белками, т.е. лишь частично включенными в фосфолипидный бислой мембраны, о чем можно судить по их спонтанной солюбилизации (рис. 1). По-видимому, в естественных условиях мембранные ферменты существуют в виде олигомеров, примером чему служит сахаразно-изомальтазный комплекс.
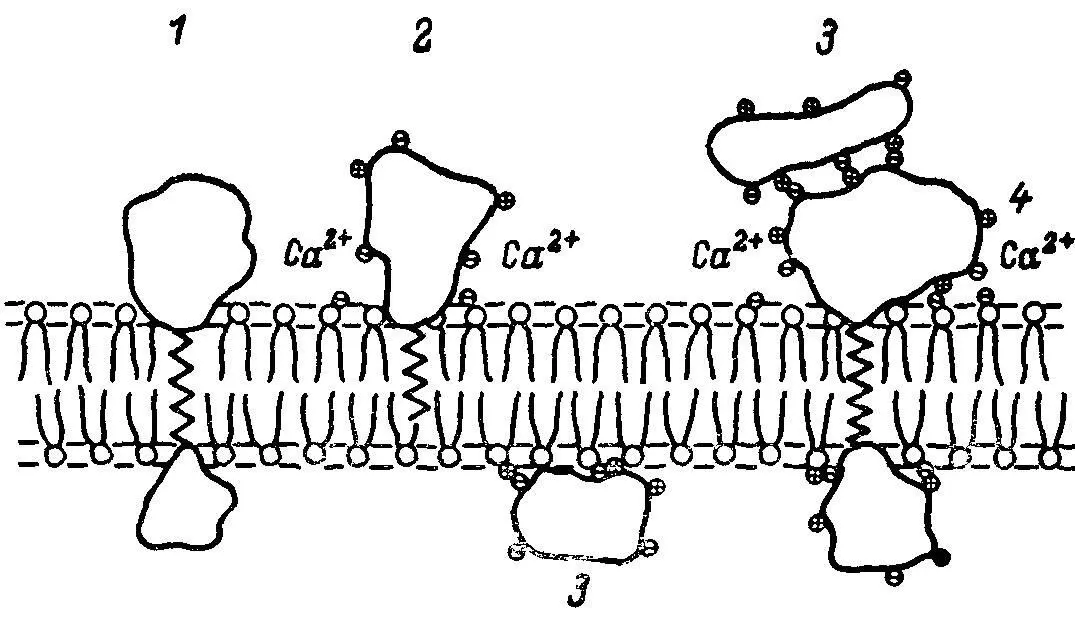
Рис. 1. Схема различных типов локализации интегральных белков в мембране.
1,4 — трансмембранные интегральные белки; 2 — периферический интегральный белок; 3 — периферический белок.
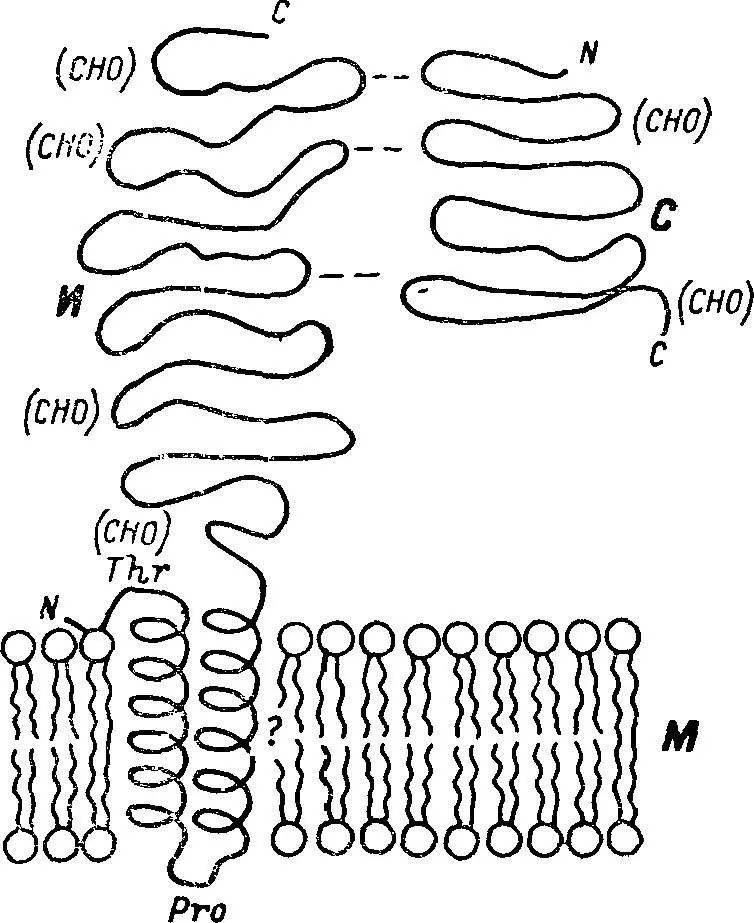
Рис. 2. Положение сахаразно-изомальтазного комплекса в щеточнокаемной мембране клеток тонкой кишки.
(СНО) — углеводные цепи; с — С-конец цепи; N—N-конец цепи; С — сахаразная субъединица; И — изомальтазная субъединица; М — мембрана.
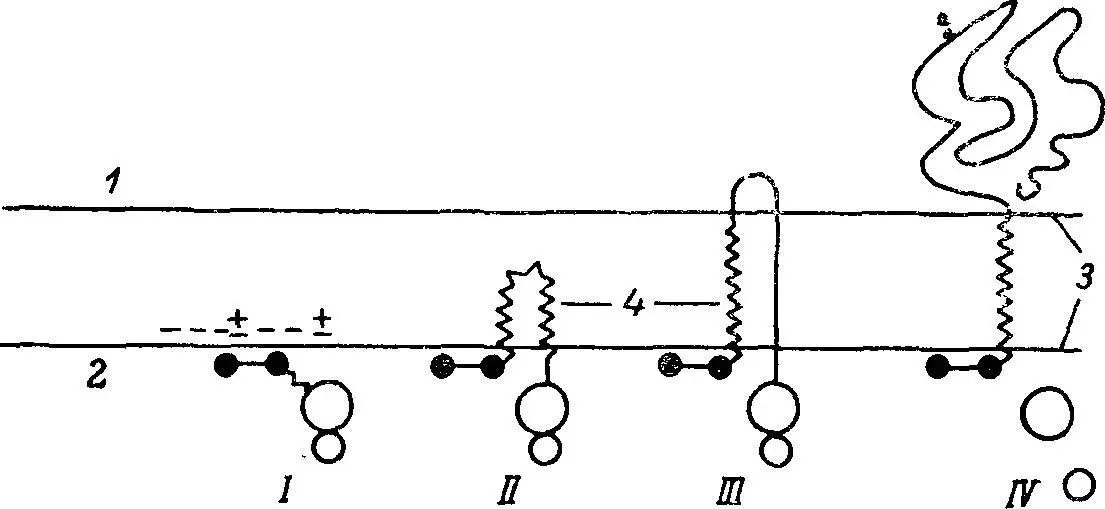
Рис. 3. Модель включения кишечной щеточнокаемной аминопептидазы в мембрану за счет N-концевых аминокислотных остатков.
1 — внеплазматическая поверхность; 2 — цитоплазматическая поверхность; 3 —мембрана; 4 — гидрофобный домен фермента, состоящий примерно из 20 аминокислотных остатков; «+» — положительно заряженный N-концевой тирозин и лизин в позиции 4; «—» — отрицательно заряженные внутренние фосфолипидные головки (или цвиттерионы). I — связывание образующейся цепи благодаря электростатическим взаимодействиям с цитоплазматической поверхностью мембраны; II — прямое перемещение гидрофобной части фермента во внутреннюю область мембраны; III, IV — дальнейший рост цепи за счет предполагаемого механизма, выдвигающего цепь на внеплазматическую поверхность мембраны.
Часть фермента может выступать над поверхностью трехслойной мембраны примерно на 10—15 нм. Простетическая группа молекулы фермента — олигосахаридная цепь (цепи) занимает наружное положение по отношению к мембране и выдается в полость тонкой кишки, возможно, принимая участие в формировании структур гликокаликса, рецепции биологически активных веществ и связывании субстратов (рис. 2).
Мембранные интегральные ферменты обладают амфипатической структурой и состоят из гидрофильного и гидрофобного доменов. Гидрофильный домен, составляющий от 90 до 95% массы фермента, несет на себе углеводные остатки и каталитический центр, обращенный в полость тонкой кишки. Такая локализация каталитического центра постулирована нами еще в начале 60-х гг. и окончательно подтверждена в последние годы. Остальная масса фермента относится к гидрофобному домену, который состоит преимущественно из гидрофобных аминокислот и пронизывает насквозь фосфолипидный бислой, в некоторых случаях заканчиваясь небольшим гидрофильным пептидом, экспонированным на внутренней поверхности мембраны (рис. 3). По мнению ряда исследователей, роль гидрофобного домена фермента заключается во взаимодействии с фосфолипидным матриксом мембраны и в фиксации гидрофильного каталитического домена (якорная функция).
Читать дальшеИнтервал:
Закладка:









