Марк Мосевицкий - Распространненость жизни и уникальность разума?

- Название:Распространненость жизни и уникальность разума?
- Автор:
- Жанр:
- Издательство:ПИЯФ РАН
- Год:2008
- Город:Санкт-Петербург
- ISBN:5-86763-203-2
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Марк Мосевицкий - Распространненость жизни и уникальность разума? краткое содержание
Исследование самых ранних осадочных пород показывает, что клеточные формы жизни присутствовали на Земле около 3.85 млрд лет тому назад, т. е. появились вскоре после создания совместимых с жизнью условий. Это означает, что доклеточная эволюция при всей ее сложности могла быть осуществлена в очень короткий срок (10–50 млн лет). Согласно другой версии, доклеточная эволюция происходила вне Земли (панспермия). В любом случае пример Земли позволяет предполагать широкое распространение жизни везде, где созданы подходящие для ее развития условия. С другой стороны, палеонтологическая летопись, начатая около 3.5 млрд лет тому назад, позволила выявить многочисленные кризисы (вымирания), приводившие к крутым разворотам в ходе эволюционного процесса. В основе эволюционных кризисов лежали природные катастрофы, вызванные падениями астероидов, глобальными оледенениями, длительными (в течение миллионов лет) истечениями лавы, изменениями газового баланса в океане и атмосфере. Бесчисленные воздействия на эволюцию оказывали также происходившие спонтанно мутации и горизонтальные обмены генетическим материалом. Благодаря наложению большого числа случайных факторов эволюционный процесс каждый раз является уникальным. На Земле после почти 4-х млрд лет существования жизни эволюционный процесс дал единственную ведущую к разуму ветвь гоминин (прямоходящих наследников шимпанзе). После многих критических ситуаций на этой ветви сохранился один побег – современное человечество. Отсутствие признаков существования других цивилизаций в доступном для общения космосе подтверждает весьма низкую вероятность выхода эволюции на маршрут, ведущий к разуму. Понимание природы кризисных событий, имевших место на Земле в прошлом, необходимо для того, чтобы, опираясь на все возрастающие возможности человечества, предотвратить наступление этих событий в будущем или хотя бы смягчить их последствия. Основной материал доступен широкому кругу читателей. Автор стремился также сделать книгу интересной для специалистов. Этим, в частности, объясняется большое число ссылок на последние публикации в научных журналах.
Распространненость жизни и уникальность разума? - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
В образовании химических связей между аминокислотами, удерживаемыми в петлях РНК-матрицы, могли участвовать как РНК-ферменты (рибозимы), так и ферменты белковой природы. Этот вопрос остается открытым. Однако в современном аппарате трансляции на рибосомах этап образования пептидной связи осуществляется с участием элемента рибосомной РНК в качестве рибозима (Joyce, 1989; Lhose and Szostak, 1996; Zhang and Cech, 1997).
Химические модификации азотистых оснований, а также ошибки при копировании (подключение к растущей цепи не комплементарного матрице азотистого основания) являлись причиной мутаций в РНК-матрицах, которые приводили к конформационным изменениям петель и обеспечивали их изменчивость как адапторов аминокислот. Соответственно эволюционировали контролируемые РНК-матрицами белки. Другой кардинальный путь модификации РНК-матриц, имеющий следствием появление новых белков, – рекомбинация матриц как путем прямого обмена материалом, так и по механизму перемены матрицы при репликации (Kogoma, 1996).
Необходимым элементом рассматриваемого механизма примитивного кодирования белков является плотная укладка РНК-матрицы, обеспечивающая ее компактность и тесное прилегание соседних петель друг к другу (как монеты в стопке). При должном ориентировании аминокислот, связанных с соседними петлями, их концевые амино и карбоксильная группы оказывались достаточно сближенными для образования химической связи. Можно предположить, что определенная конформация матричной молекулы РНК, обеспечивавшая координированную позицию петель и их тесную упаковку, была стабилизирована взаимодействием боковых участков соседних петель. Интересно отметить, что, хотя в современной системе трансляции основную стабилизирующую функцию выполняет рибосома, взаимодействие петлевых элементов двух молекул тРНК, которые доставили в рибосому очередные аминокислоты, сохраняет значение как дополнительный стабилизирующий фактор (Smith and Yarus, 1989).
3.2. Переход к современному способу кодирования аминокислотных последовательностей в белках
Мир РНК явился переломным в эволюции клетки, т. к. в рамках этого мира произошел переход от образования пептидов на организованных в форме петель матрицах к современному способу кодирования наследственной информации и отделенному от этого процесса синтезу белков в сложно организованных РНК-белковых структурах – рибосомах. В этом синтезе, помимо рибосомных РНК, участвуют информационные (messendger) РНК (мРНК), в нуклеотидных последовательностях которых закодированы аминокислотные последовательности белков, и адапторные или транспортные РНК (тРНК), которые связываются с определенными аминокислотами и доставляют их в рибосомы к экспонированным кодонам мРНК. Одной из интригующих загадок эволюции является вопрос, появились мРНК и тРНК согласованно как “запланированные” элементы системы кодирования белков или возникли независимо, не будучи исходно предназначены для целей кодирования. До настоящего времени общепринятого решения этой проблемы нет. Автору представляется предпочтительной гипотеза “нецелевого” (по Дарвину) образования этих типов РНК, которые впоследствии оказались тесно взаимодействующими элементами новой системы кодирования и синтеза белков.
Имея в виду эту гипотезу, еще раз обратим внимание на то обстоятельство, что петли РНК-матрицы были тесно сближены. Это могло привести в действие процедуру репликации (копирования) со сменой матрицы (copy choice) (Kogoma, 1996): сближенные участки петель, которые мы достаточно условно именуем торцевыми, служили элементами прерывистой матрицы, по которой шел синтез непрерывной комплементарной копии (Рис. 1Б). Нуклеотидную последовательность этой “огибающей” нити можно условно разбить на ряд коротких участков, которые соответствовали определенным петлям (были комплементарны их торцевым участкам) и следовали в том же порядке. Очевидно, что благодаря такой структуре, возникшая первоначально как побочный продукт “огибающая” нить несла информацию о последовательности аминокислот в пептиде. Эта нить, как и другие молекулы РНК, подвергалась авторепликации, благодаря чему зафиксированная в ней информация оказывалась тиражирована в потомстве. Однако в условиях примитивного кодирования белков информационные возможности огибающих РНК оставались невостребованными. Первоначально они могли выполнять роль дополнительного стабилизатора конформации РНК-матрицы с тесно сближенными петлями, осуществляя с их торцами комплементарное взаимодействие. Однако в клетках наряду с полными РНК-матрицами присутствовали их фрагменты, возникавшие как при прерывании синтеза РНК-матриц, так и при деградации полных молекул. С помощью огибающей РНК фрагменты РНК-матрицы могли быть выстроены в “правильной” последовательности, образуя в сумме полную матрицу.
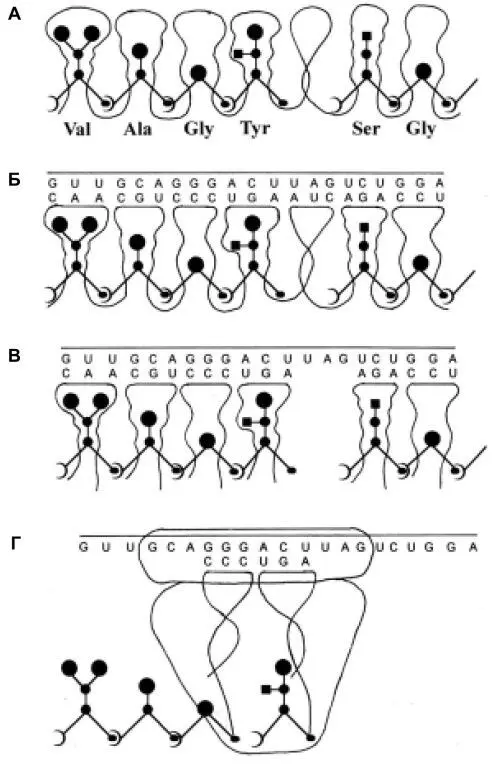
Рис. 1.
Рис. 1. Схемы, иллюстрирующие гипотезу перехода от примитивного синтеза предетерминированных пептидов к современному генетическому коду.
А. Образование белка (пептида), запрограммированного последовательностью петель-полостей единой молекулы РНК, связывающих активированные аминокислоты. Б. То же. Показана также “огибающая” РНК, нуклеотидная последовательность которой соответствует (комплементарна) ряду “торцов” петлевых элементов матрицы. В. Синтез пептидов на последовательности разделенных полостей-петель, набираемых из общего пула. Порядок петлевых элементов в наборе определяется связыванием их торцевых участков с соответствующими участками «огибающей» РНК, выполняющей таким образом роль информационной молекулы (мРНК). Соответственно, петлевые элементы в этой схеме являются предшественниками тРНК. Г. Эпизод современного синтеза белка на рибосоме: присоединение очередного звена к растущей белковой цепи. Вследствие изменения механизма узнавания адапторами соответствующих аминокислот петли тРНК перестали представлять собою «полости», хотя в них присутствуют необходимые для узнавания структурные элементы.
Схемы Рис. 1 предназначены для иллюстрации основных положений гипотезы и не отражают истинной конформации (укладки) матричных молекул РНК. Другие разъяснения приведены в тексте. Рисунок создан при участии В.В. Горбенко.
И это уже можно рассматривать как частичную реализацию информационных возможностей огибающих нитей. До полной реализации этих возможностей оставался один шаг: образование пула автореплицирующихся автономных РНК-адапторов, связывавших отдельные аминокислоты и узнававших соответствующие им участки в информационных нитях. Первоначально адапторные молекулы – предшественницы современных транспортных РНК (тРНК) – возникали как однопетлевые фрагменты РНК-матрицы. Сформировавшийся пул автономных РНК-петель обеспечивал связывание всех аминокислот, входивших тогда в состав пептидов. “Заряженные” аминокислотами автономные петли матричной РНК были также способны своими торцевыми участками подключаться к соответствующим (комплементарным) участкам огибающей РНК. Принципиально важным для дальнейшей эволюции качеством пула автономных адапторов аминокислот стала его универсальность: он мог быть использован при синтезе любого пептида в присутствии соответствующей огибающей РНК.
Читать дальшеИнтервал:
Закладка:







