Марк Мосевицкий - Распространненость жизни и уникальность разума?

- Название:Распространненость жизни и уникальность разума?
- Автор:
- Жанр:
- Издательство:ПИЯФ РАН
- Год:2008
- Город:Санкт-Петербург
- ISBN:5-86763-203-2
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Марк Мосевицкий - Распространненость жизни и уникальность разума? краткое содержание
Исследование самых ранних осадочных пород показывает, что клеточные формы жизни присутствовали на Земле около 3.85 млрд лет тому назад, т. е. появились вскоре после создания совместимых с жизнью условий. Это означает, что доклеточная эволюция при всей ее сложности могла быть осуществлена в очень короткий срок (10–50 млн лет). Согласно другой версии, доклеточная эволюция происходила вне Земли (панспермия). В любом случае пример Земли позволяет предполагать широкое распространение жизни везде, где созданы подходящие для ее развития условия. С другой стороны, палеонтологическая летопись, начатая около 3.5 млрд лет тому назад, позволила выявить многочисленные кризисы (вымирания), приводившие к крутым разворотам в ходе эволюционного процесса. В основе эволюционных кризисов лежали природные катастрофы, вызванные падениями астероидов, глобальными оледенениями, длительными (в течение миллионов лет) истечениями лавы, изменениями газового баланса в океане и атмосфере. Бесчисленные воздействия на эволюцию оказывали также происходившие спонтанно мутации и горизонтальные обмены генетическим материалом. Благодаря наложению большого числа случайных факторов эволюционный процесс каждый раз является уникальным. На Земле после почти 4-х млрд лет существования жизни эволюционный процесс дал единственную ведущую к разуму ветвь гоминин (прямоходящих наследников шимпанзе). После многих критических ситуаций на этой ветви сохранился один побег – современное человечество. Отсутствие признаков существования других цивилизаций в доступном для общения космосе подтверждает весьма низкую вероятность выхода эволюции на маршрут, ведущий к разуму. Понимание природы кризисных событий, имевших место на Земле в прошлом, необходимо для того, чтобы, опираясь на все возрастающие возможности человечества, предотвратить наступление этих событий в будущем или хотя бы смягчить их последствия. Основной материал доступен широкому кругу читателей. Автор стремился также сделать книгу интересной для специалистов. Этим, в частности, объясняется большое число ссылок на последние публикации в научных журналах.
Распространненость жизни и уникальность разума? - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
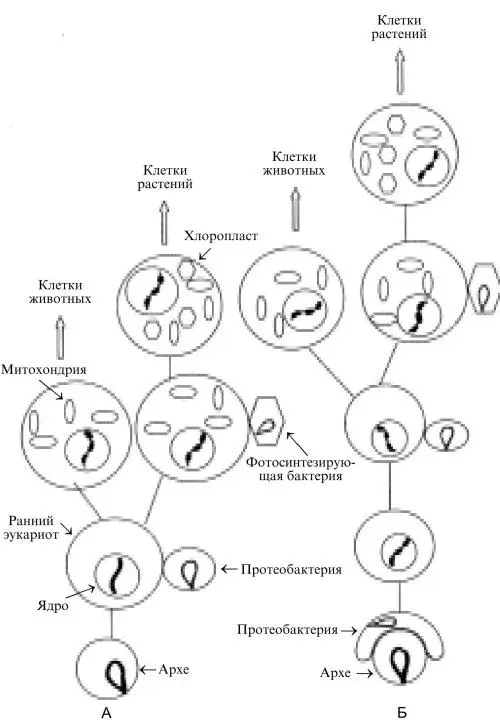
Рис. 5. Формирование современных эукариотических клеток.
А. На базе возникших эволюционным путем ранних эукариотов. Б. На базе эукариота, возникшего благодаря акту симбиоза протеобактерии и клетки архе (подробности в тексте). Рисунок создан при участии О. М. Горбенко.
Однако авторы, поддерживающие эту концепцию, расходятся в предположении о принадлежности бактерии, участвовавшей в симбиозе с архе, той или иной группе протеобактерий. Это свидетельствует о необходимости более детальных исследований. Тот факт, что какие-то гены эукариот происходят от архе, а другие – от бактерий, позволяет объяснить, почему авторы, использовавшие ранее филогенетический (мутационный) подход, вели происхождение эукариот как от архе (Woese et al., 1990; Doolittle, 1999), так и от бактерий (Hashimoto et al., 1998): все дело в том, ген какого происхождения (архе или бактериального) был выбран для исследования. Cформировавшиеся указанным образом эукариоты позже приобрели митохондрии и хлоропласты благодаря последовательным актам симбиоза с α-протеобактерией и с фотосинтезирующими грамотрицательными бактериями, о чем уже упоминалось выше.
В рамках описанной гипотезы уместно поставить вопрос, почему слияние двух генетически различных прокариотических клеток стало стимулом для формирования клеточного ядра, т. е. образования эукариота. Интересную гипотезу предлагают Мартин и Кунин (Martin and Koonin, 2006). Эти авторы обращают внимание на то обстоятельство, что при формировании в симбионте объединенных геномов происходила фрагментация некоторых генов вследствие образования чужеродных вставок. Однако трансляция непосредственного транскрипта разобщенного гена давала “неправильный” продукт. Для получения прежнего белка следовало освободить мРНК от вставок (интронов). Необходимый для этого процесс вырезания интронов и сшивки фрагментов мРНК (экзонов), именуемый сплайсингом, должен предшествовать трансляции (образованию белков на рибосомах). Предотвратить трансляцию первичных транскриптов можно было, пространственно разделив оба процесса. Именно эту цель преследовало формирование внутренней (ядерной) оболочки. Она изолировала хромосомы от остального объема клетки (цитоплазмы), куда переместились рибосомы, происходившие от клетки архе, занимавшей центральную область симбионта. Образованные в ядре транскрипты после достаточно изощренной и, кстати, небыстрой процедуры сплайсинга уже в форме кодирующих нормальный продукт мРНК транспортируются через ядерные поры в цитоплазму, где подвергаются трансляции на располагающихся там рибосомах. На некоторые данные, вызывающие сомнения в правомерности этой гипотезы, указывают сами авторы, отмечая периодически появляющиеся публикации об осуществлении трансляции в самом ядре на присутствующих там “ядерных” рибосомах (Iborra et al., 2001). Однако утверждение о присутствии рибосом в ядре нельзя считать доказанным (см. Dahlberg and Lund, 2004). Можно высказать и другое соображение. Процессы эволюционного создания инструментов сплайсинга, формирования изолированного от цитоплазмы ядра и отделения рибосом от первичных транскриптов могли занять сотни и сотни поколений. Вместе с тем, клетки для поддержания жизнеспособности должны постоянно синтезировать белки, обладающие необходимой активностью. Это вряд ли выполнимо в случае трансляции первичных транскриптов, содержащих интроны, что неизбежно приводило бы к образованию белков с измененной структурой и нарушенными свойствами. Эту проблему, которая кажется трудноразрешимой, авторы не обсуждают. Авторы рассмотренных выше гипотез формирования эукариотической клетки из композитной клетки, образованной слиянием архе и бактерии, оставляют без ответа и ряд других вопросов. Именно это послужило причиной появления новых идей.
Значительное количество эукариотических генов не имеет аналогов ни у бактерий, ни у архе. Хартман и Федоров (Hartman and Fedorov, 2002) выявили в эукариотических организмах (дрожжах, дрозофиле и др.) более 300 белков, не имеющих “родственников” у известных прокариот. Основываясь на этом факте, авторы предположили участие в формировании эукариотической клетки еще и третьего участника, также прокариота, принадлежавшего линии, названной ими хроноцитами. Эта линия сама не сохранилась и оставила след только в сформированных с ее участием эукариотах. Имея в виду функции белков, отсутствовавших у бактерий и у архе, Хартман и Федоров предполагают, что главной особенностью хроноцитов было присутствие цитоскелета и других структурных элементов цитоплазмы (шероховатый ретикулум, тельца Гольджи), которые отличают цитоплазму всех эукариот от почти неструктурированной цитоплазмы известных прокариот. Как полагают эти авторы, ядра сформировались в результате эндосимбиотического взаимодействия бактерии и архе в хроноците. Последний, уже обладавший актиновым и тубулиновым цитоскелетом, предоставил генетическую информацию, составившую базу для формирования сложно организованной цитоплазмы эукариотической клетки.
Однако было ли действительно необходимым привнесение в композит гипотетическим хроноцитом определенных признаков, которые отсутствовали у двух очевидных симбионтов – бактерий и архе? В принципе, такой необходимости не было, так как значительное количество новых признаков (генов) сформировалось в процессе эволюционного развития эукариота, т. е. уже после образования композита. В первую очередь это относится к самому ядру. О необходимости формирования у эукариот новых механизмов (а следовательно, и генов) в связи с организацией сплайсинга говорилось выше. Вообще, клеточное ядро – весьма сложное образование. Помимо двуслойной ядерной оболочки, пронизанной регулярно распределенными белковыми структурами, образующими ядерные поры, клеточное ядро имеет сложное внутреннее строение. Его фибриллярный скелет (ядерный матрикс) образован, главным образом, ядерными белками ламинами. Детали структуры ядерного матрикса на протяжении многих лет являются предметом обсуждения (см. Георгиев и Ченцов, 1960; Berezney and Coffey, 1977; Gerace and Blobel, 1982; Мосевицкий и Новицкая, 1982; Мосевицкий, 1985; van Eekelen et al., 2002). Ядерный матрикс участвует в организации и функционировании заключенных в ядре хромосом. На нем базируются ферментные комплексы, осуществляющие пооперонную репликацию ДНК (Tubo and Berezney 1987), транскрипцию (Razin et al., 1985; Jackson, 2005), сплайсинг (Ciejek et al., 1982; Mariman et al., 1982) и другие функции (Pienta and Coffey, 1984). Следует добавить также специфические для эукариот белки, обеспечивающие компактную форму хроматина (гистоны), системы, контролирующие регулярность деления эукариотической клетки (митоз) и др. Мы видим, что объем генетической информации, которую необходимо было освоить формировавшемуся эукариоту (в форме новых генов), был очень велик. Создание цитоскелета стало также необходимым условием образования современной эукариотической клетки, так как позволяло увеличить клеточный объем, фиксировать положение ядра, перейти к активному транспорту и др. Но и эта задача могла решаться уже после акта симбиоза, инициировавшего весь процесс формирования эукариотической клетки. Следует также иметь в виду, что такой акт мог бы состояться 3 и даже 3.5 млрд лет тому назад, когда потенциальные симбионты (архе и соответствующие формы бактерий) уже существовали на Земле. С другой стороны, отпечатки клеток с ядром и другими признаками, характерными для эукариотов, обнаружены в отложениях, возраст которых не более 1.7 млрд лет. Это означает, что на эволюцию от момента симбиотического образования химерной клетки до появления эукариотической клетки было отпущено свыше одного миллиарда лет – даже больше, чем на всю предшествовавшую эволюцию на Земле. Сколько на самом деле продолжался этот процесс – неизвестно, т. к. неясно и вряд ли может быть установлено, когда состоялся инициировавший его акт симбиоза.
Читать дальшеИнтервал:
Закладка:







