Евгений Кунин - Логика случая. О природе и происхождении биологической эволюции

- Название:Логика случая. О природе и происхождении биологической эволюции
- Автор:
- Жанр:
- Издательство:Литагент «Центрполиграф»a8b439f2-3900-11e0-8c7e-ec5afce481d9
- Год:2014
- Город:Москва
- ISBN:978-5-227-04982-7
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Евгений Кунин - Логика случая. О природе и происхождении биологической эволюции краткое содержание
В этой амбициозной книге Евгений Кунин освещает переплетение случайного и закономерного, лежащих в основе самой сути жизни. В попытке достичь более глубокого понимания взаимного влияния случайности и необходимости, двигающих вперед биологическую эволюцию, Кунин сводит воедино новые данные и концепции, намечая при этом дорогу, ведущую за пределы синтетической теории эво люции. Он интерпретирует эволюцию как стохастический процесс, основанный на заранее непредвиденных обстоятельствах, ограниченный необходимостью поддержки клеточной организации и направляемый процессом адаптации. Для поддержки своих выводов он объединяет между собой множество концептуальных идей: сравнительную геномику, проливающую свет на предковые формы; новое понимание шаблонов, способов и непредсказуемости процесса эволюции; достижения в изучении экспрессии генов, распространенности белков и других фенотипических молекулярных характеристик; применение методов статистической физики для изучения генов и геномов и новый взгляд на вероятность самопроизвольного появления жизни, порождаемый современной космологией.
Логика случая демонстрирует, что то понимание эволюции, которое было выработано наукой XX века, является устаревшим и неполным, и обрисовывает фундаментально новый подход – вызывающий, иногда противоречивый, но всегда основанный на твердых научных знаниях.
Логика случая. О природе и происхождении биологической эволюции - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Чем же тогда объяснить неожиданно слабую связь между эволюцией и функцией гена? По отношению к эволюции последовательностей можно предположить, что скорость эволюции более зависит от внутренних характеристик гена (в особенности от структуры кодируемого белка), чем от его биологической значимости. Однако это объяснение неприменимо к случаям утраты гена. Наиболее осмысленным – хотя опять-таки противоречащим здравому смыслу – кажется следующее объяснение: фенотипический эффект нокаута гена (и в целом – набора необходимых генов) не проявляется как консервативное свойство в эволюционном процессе и быстро меняется (в масштабах эволюционной шкалы), вероятно благодаря высокой скорости эволюции сетей взаимодействующих генов [39]. Ясно, что это предположение можно проверить опытным путем, пусть и с помощью трудоемких экспериментов.
Почти нейтральные сети и белковая эволюция
В целом скорость эволюции гена определяется размером его почти нейтральной сети, то есть множества последовательностей, получаемых друг из друга в результате одношаговых мутаций (пусть и необязательно с помощью одной замены) и имеющих приспособленность примерно такую же, как и у наиболее приспособленной последовательности (Wagner, 2008a; Wolf et al., 2010). Чем больше нейтральная сеть, тем слабее ограничения для конкретного гена, тем быстрее он может эволюционировать (рис. 4–5).
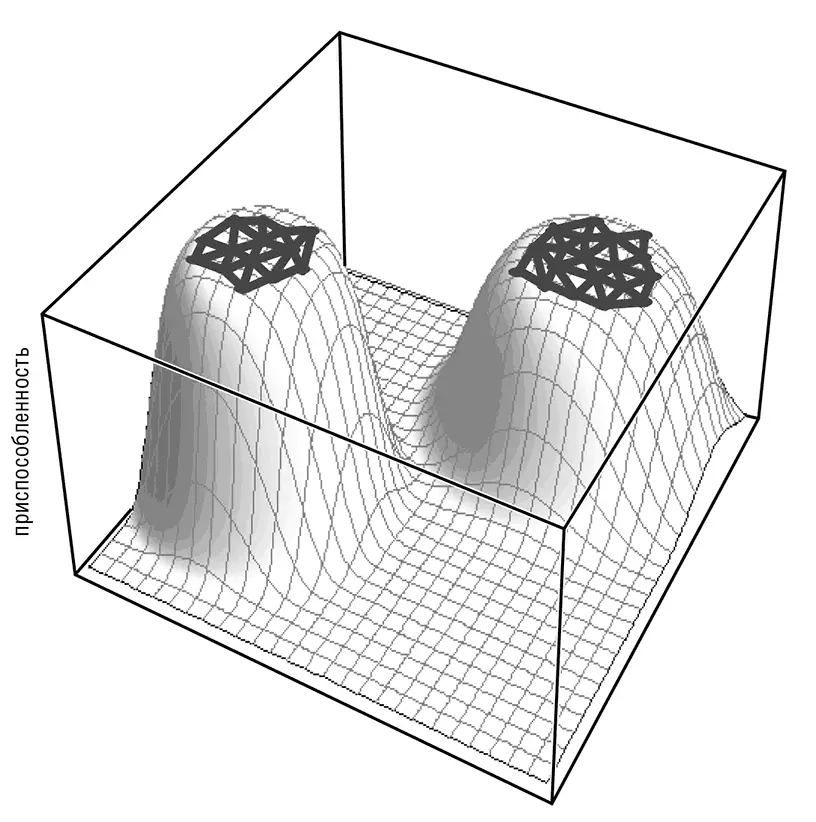
Рис. 4–5. Почти нейтральная сеть и белковая эволюция. Две почти нейтральные сети для двух вымышленных белков схематично представлены как две области, находящиеся на широких вершинах пиков приспособленности.
В эволюции белка приспособленность отдельной последовательности в основном зависит от ее устойчивости к ошибкам укладки и от уровня экспрессии, а размер почти нейтральной сети зависит от высоты и формы пика, занимаемого этой последовательностью и ее соседями на ландшафте устойчивости (рис. 4–6). В этой модели высокоэкспрессированные белки, чьи исходные последовательности высоко устойчивы к ошибкам укладки, занимают высокие и крутые пики с небольшой областью высокой приспособленности (малые почти нейтральные сети) и, следовательно, подвержены сильному стабилизирующему отбору и медленно эволюционируют. И наоборот, белки с более низким уровнем экспрессии и меньшей устойчивостью находятся на более низких, пологих пиках, имеют более широкую область высокой приспособленности и, соответственно, подвержены более слабому отбору и имеют высокую скорость эволюции (рис. 4–6; Wolf et al., 2010).
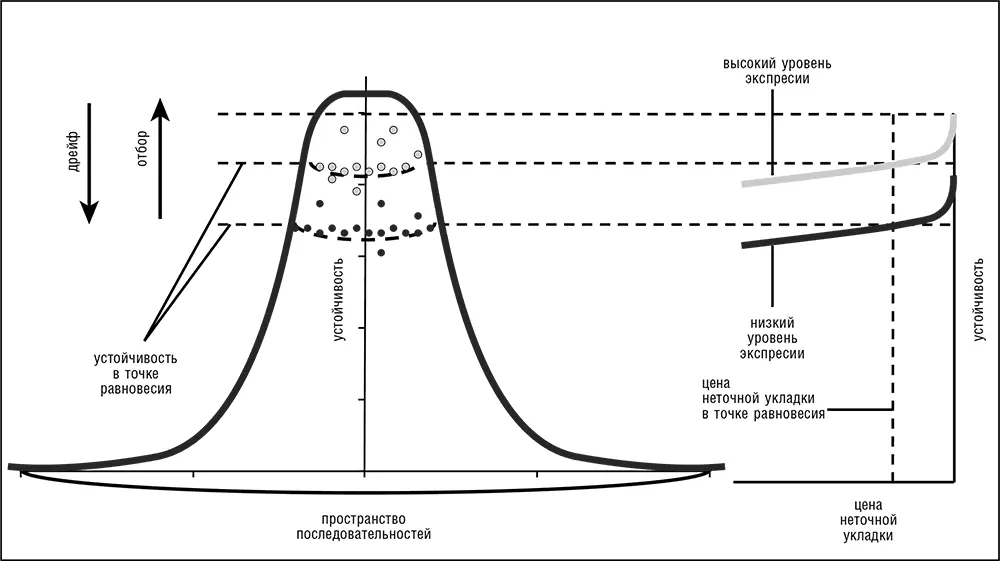
Рис. 4–6. Модель эволюции белка, управляемой издержками неточной укладки.
Геномная эволюция путем дупликации генов, модель рождения и смерти гена и универсальное распределение численности паралогичных семейств
Мы уже касались вопроса дупликации генов в предыдущей главе, в частности при построении списка важнейших механизмов геномной эволюции. Однако есть еще по крайней мере два хороших повода вернуться к этому механизму эволюции и обсудить его более детально. Во-первых, дупликация несомненно является одним из главных путей геномной эволюции для всех форм жизни и играет принципиальную роль в эволюции эукариот (см. гл. 8). Во-вторых, эволюция путем дупликации генов представляет собой формально простой процесс, для которого довольно легко построить хорошо работающие физические (или математические) модели, которые и рассматриваются в этой главе.
Представление о дупликации как об исключительно эффективном способе геномной эволюции лежит в основе современного эволюционного мышления. Упрощенно говоря, сущность этого представления состоит в том, что создание новых функциональных объектов (белков и РНК) путем модификации уже имеющихся (вспомните модель «эволюции как мастерового» Жакоба; Jacob, 1977) – это, очевидно, намного более простой путь, чем создание этих объектов с самого начала, с нуля (история этой идеи рассказывается в гл. 2). Как и другие представления геномики, эволюция путем генной дупликации имеет строгие количественные подтверждения – большинство генов в геноме принадлежат семействам паралогов (за исключением очень небольших геномов, таких как у микоплазмы и других паразитических бактерий; Jordan et al., 2001). Более детальная реконструкция эволюции показывает, что дупликация проявляется, с различной интенсивностью, на всех этапах эволюции, таким образом, любой геном – это набор дупликаций самого различного возраста. Выбирая некоторую эволюционную линию, допустим, животные – хордовые – млекопитающие – приматы – и т. д., мы можем обнаружить в геноме (например, в нашем) все соответствующие классы дупликаций: дупликации, специфичные для животных, дупликации, специфичные для хордовых, специфичные для приматов и т. д. (Lespinet et al., 2002).
Распределение численности паралогичных семейств в любом геноме – еще одна универсальная статистическая закономерность, обнаруженная сравнительной геномикой (рис. 4–7). Распределения для всех геномов приблизительно описываются степенной функцией с отрицательным показателем степени: y = ах -y (где у– положительное число, а– коэффициент; Koonin et al., 2002; Luscombe et al., 2002). Эти распределения, имеющие в двойных логарифмических координатах вид прямых линий, показывают, что большинство семейств по численности малы (включая семейства геномов с преобладанием синглетонных сайтов), и только немногие семейства включают в себя большое число паралогов.
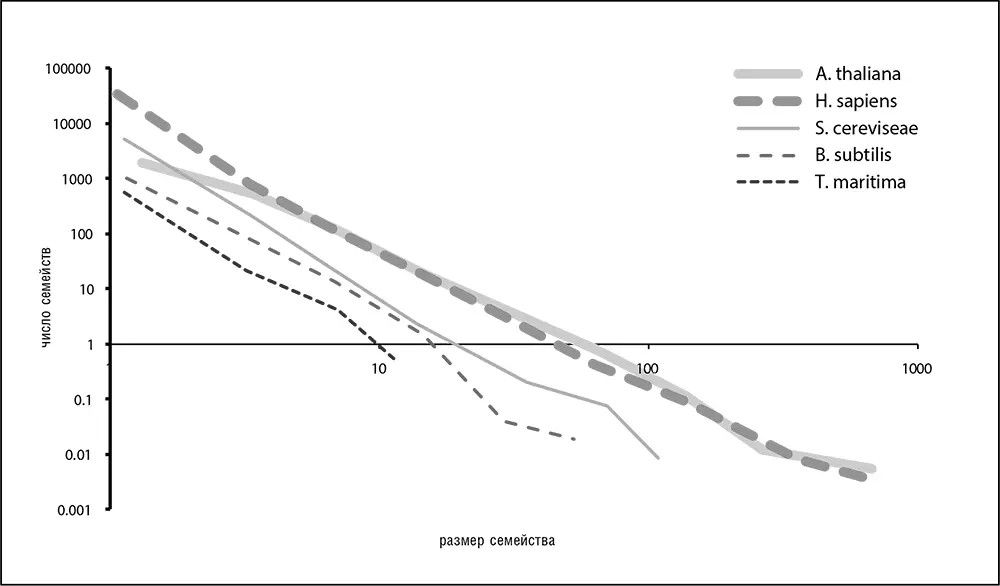
Рис. 4–7. Распределение размера семейств паралогичных генов для нескольких сильно отличающихся геномов. Показаны распределения для растения резуховидки Таля ( A. thaliana ), человека ( H. sapiens ), для пекарских дрожжей ( S. cereviseae ), для сенной палочки ( B. subtilis ) и для бактерии Thermotoga maritima . Использованы данные из базы данных EggNog (Jensen et al., 2008)
Возникновение универсальной степенной закономерности распределения численности паралогичных семейств может быть описано с высокой точностью простой математической моделью эволюционного процесса (рис. 4–8). Эта модель основана на математической теории так называемых процессов рождения и смерти (один из видов марковских процессов) и для случая эволюции путем генной дупликации чаще называется моделью рождения, смерти и инновации (Karev et al., 2002). В рамках этой модели рождение – это такая генная дупликация, при которой появляется новый член паралогичного семейства, смерть – утрата гена, а инновация – это рождение нового семейства либо путем такой дупликации, которая вызывает быструю эволюции и тем самым как бы стирает «память» старого семейства, либо путем горизонтального переноса генов [40]. Наиболее интересный результат этого моделирования состоит в том, что эта модель эволюции путем генной дупликации воспроизводит рассмотренные нами распределения численности семейств паралогичных генов только при соблюдении вполне определенных условий: частоты рождения и смерти гена должны быть примерно равными и зависеть от численности семейства таким образом, чтобы большие семейства оказывались более динамичными, чем маленькие.
Читать дальшеИнтервал:
Закладка:








