Евгений Кунин - Логика случая. О природе и происхождении биологической эволюции

- Название:Логика случая. О природе и происхождении биологической эволюции
- Автор:
- Жанр:
- Издательство:Литагент «Центрполиграф»a8b439f2-3900-11e0-8c7e-ec5afce481d9
- Год:2014
- Город:Москва
- ISBN:978-5-227-04982-7
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Евгений Кунин - Логика случая. О природе и происхождении биологической эволюции краткое содержание
В этой амбициозной книге Евгений Кунин освещает переплетение случайного и закономерного, лежащих в основе самой сути жизни. В попытке достичь более глубокого понимания взаимного влияния случайности и необходимости, двигающих вперед биологическую эволюцию, Кунин сводит воедино новые данные и концепции, намечая при этом дорогу, ведущую за пределы синтетической теории эво люции. Он интерпретирует эволюцию как стохастический процесс, основанный на заранее непредвиденных обстоятельствах, ограниченный необходимостью поддержки клеточной организации и направляемый процессом адаптации. Для поддержки своих выводов он объединяет между собой множество концептуальных идей: сравнительную геномику, проливающую свет на предковые формы; новое понимание шаблонов, способов и непредсказуемости процесса эволюции; достижения в изучении экспрессии генов, распространенности белков и других фенотипических молекулярных характеристик; применение методов статистической физики для изучения генов и геномов и новый взгляд на вероятность самопроизвольного появления жизни, порождаемый современной космологией.
Логика случая демонстрирует, что то понимание эволюции, которое было выработано наукой XX века, является устаревшим и неполным, и обрисовывает фундаментально новый подход – вызывающий, иногда противоречивый, но всегда основанный на твердых научных знаниях.
Логика случая. О природе и происхождении биологической эволюции - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
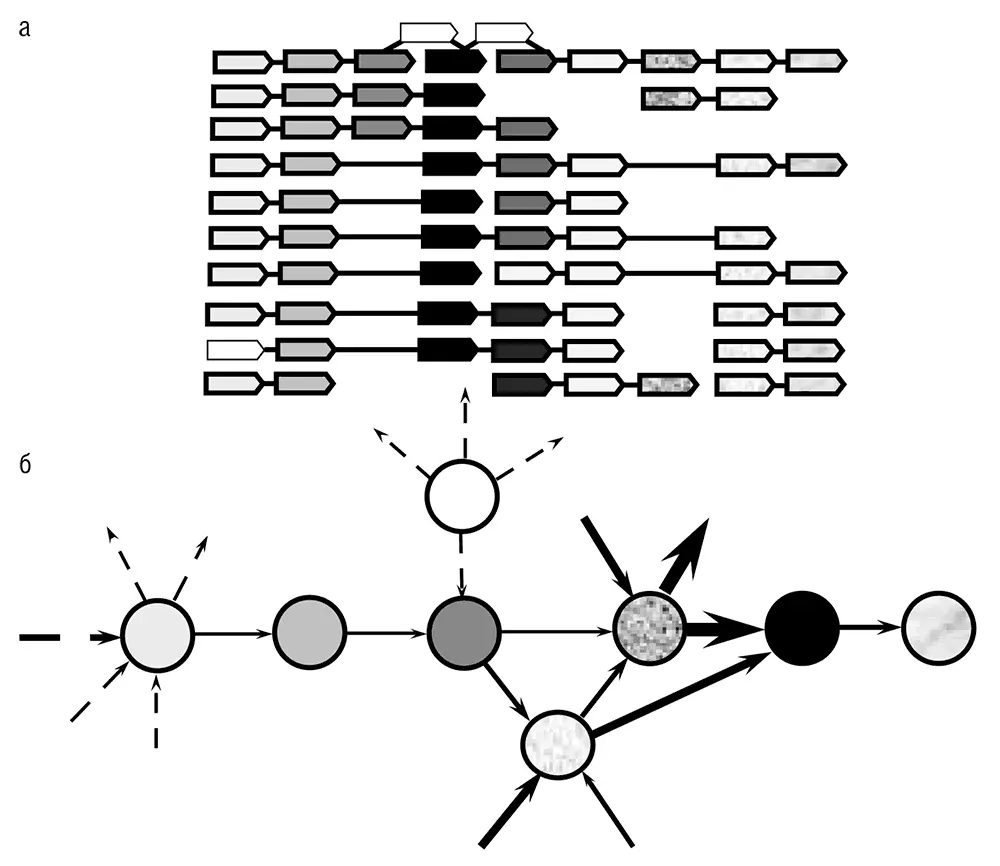
Рис. 5–3. Частично сохраняющееся окружение гена в геноме прокариот: а – перекрывающиеся, частично сохраняющиеся массивы генов. Гены показаны в форме стрелок с уникальной штриховкой или текстурой. Связывающие их жирные линии обозначают короткие межгенные промежутки, а тонкие линии показывают протяженные районы, разделяющие соответствующие гены. (Они содержат дополнительные гены и изображены без учета масштаба.) В случаях, в которых опероны не связаны, они могут располагаться в различных частях генома. На рисунке показаны реальные массивы генов, но названия конкретных геномов и генов не указаны, чтобы подчеркнуть общий характер геномной организации подобного типа. Данные по Rogozin et al., 2002; б – представление окружения гена в виде сети. Закрашенные окружности показывают гены, которые принадлежат к окружению, автоматически вычлененному с использованием алгоритма, описанного в Rogozin et al., 2002; показана только часть окружения. Незакрашенная окружность соответствует гену, который принадлежит окружению, но не был включен в него автоматической процедурой. Стрелки показывают связь между генами в оперонах (жирные стрелки соответствуют связям внутри окружения, а пунктирные стрелки – внешним связям). Толщина стрелок примерно пропорциональна числу геномов, в которых представлена данная пара генов.
Большинство оперонов находится не в сложном окружении, включающем разнообразные связи, а представляет собой простую последовательность от двух до четырех генов, порядок которых может различаться. Идентичные или похожие в смысле организации генов опероны часто обнаруживаются в сильно различающихся организмах и в различных функциональных системах. Примечательны в данном случае многочисленные опероны транспорта метаболитов, которые состоят из расположенных в одинаковом порядке генов, кодирующих трансмембранные пермеазы, АТФазы и периплазматические субъединицы так называемых ABC-транспортеров (три субъединицы обозначаются соответственно A, B и C). Присутствие таких общих оперонов в разнообразных бактериях и археях было интерпретировано в рамках гипотезы эгоистичного оперона (Lawrence, 1999), которая постулирует, что оперон так хорошо сохраняется не из-за функциональной важности совместной регуляции входящих в него генов, а из-за «эгоистичности» этой компактной генетической единицы, которая склонна к горизонтальному распространению среди прокариот (ниже в этой главе мы еще вернемся к данной концепции при обсуждении горизонтального переноса генов).
Систематическое сравнение расположения ортологичных генов в архейных и бактериальных геномах выявило относительно небольшую долю сохраняющихся (предсказанных) оперонов и гораздо большую распространенность уникальных директонов (последовательностей генов, считываемых в одинаковом направлении и разделенных короткими межгенными участками; Wolf et al., 2001). Как было показано, возможно несколько неожиданно, директоны довольно точно предсказывают опероны: большинство директонов в действительности, по-видимому, являются оперонами (Salgado et al., 2000). Таким образом, архейные и бактериальные геномы сформированы на оперонных принципах с небольшим числом высококонсервативных оперонов и намного более многочисленными редкими и уникальными оперонами. С учетом этого обстоятельства модель консервации оперонов, по крайней мере качественно, напоминает распределение кластеров ортологичных генов, с его трехкомпонентной структурой (см. выше): редкие гены и редкие опероны гораздо более многочисленны, чем повсеместно распространенные гены и опероны .
Степень «оперонизации» генома у бактерий и архей широко варьирует: некоторые геномы, например как у гипертермофильной бактерии Thermotoga maritima , почти полностью состоят из (предсказанных) оперонов, в то время как другие, как у большинства цианобактерий, по-видимому, содержат очень немного оперонов. Остается неясным, что определяет распространенность оперонов в организме, хотя высказывались предположения, что степень «оперонизации» зависит от баланса между интенсивностью рекомбинации и горизонтального потока генов, а также факторов отбора, препятствующих разрушению оперонов.
Регуляция экспрессии генов и передачи сигналов у бактерий и архей: от базовой схемы оперона к сверхоперонам, регулонам и сложным сетям
Бактерии и археи обладают сложной и элегантной системой регуляции экспрессии генов. Сравнительная геномика драматически изменила существующие взгляды на принципы организации, распределение в природе и эволюцию этих регуляторных механизмов. Концепция оперона Жакоба и Моно, представленная в предыдущем разделе как основной принцип локальной архитектуры бактериальных и архейных геномов, также является концепцией регуляции экспрессии генов и передачи сигналов у прокариот. В модели Жакоба – Моно регулятор (репрессор лактозы в их оригинальной работе) является сенсором внеклеточных и внутриклеточных сигналов (в данном случае концентрации лактозы), что влияет на структуру белка-регулятора и, опосредованно, на экспрессию оперона (в случае лактозного оперона репрессор, связывая лактозу, отсоединяется от регуляторной части оперона, делая тем самым возможной транскрипцию). В течение полувека, прошедших с момента фундаментального открытия Жакоба – Моно, было обнаружено множество вариаций этой темы, включая регуляторы, которые симметрично влияют на транскрипцию разных расположенных по соседству генов, и глобальные регуляторы, которые контролируют экспрессию многочисленных разрозненных генов и оперонов, в противоположность репрессору простого оперона в модели Жакоба – Моно. Наиболее заметными глобальными регуляторами являются белки – подавители катаболизма (CRP) и регулятор ответа на стресс (SOS) LexA. С учетом открытия этих и других глобальных регуляторов концепция оперона была усовершенствована понятием регулона – набора генов, экспрессия которых регулируется одним и тем же белком-регулятором. Сравнительный геномный анализ регулонов выявил их чрезвычайную эволюционную пластичность с существенными различиями между регулонами даже у близкородственных организмов (Lozada-Chavez et al., 2006). Глобальные регуляторы транскрипции, такие как LexA, широко распространены и высококонсервативны в различных бактериях, но состав генов в регулоне LexA является очень вариативным. Пластичность регулонов, наряду с изменчивостью архитектуры генома (см. выше), хорошо согласуется с идеей, что регуляция экспрессии генов и архитектура генома в эволюции архей и бактерий тесно взаимосвязаны. В резком контрасте с изменчивостью и пластичностью регулонов, регуляторы транскрипции у бактерий и архей демонстрируют примечательное единство архитектуры и структуры. Как правило, эти регуляторы содержат домен, связывающий небольшие молекулы-сенсоры и ДНК-связывающий домен. Подавляющее большинство ДНК-связывающих доменов являются вариациями одной и той же структурной темы, спираль – поворот – спираль. Более специфические, но тоже распространенные домены связывания с ДНК включают мотивы лента – спираль – спираль и цинковая лента (Aravind et al., 2005; Aravind and Koonin, 1999).
Читать дальшеИнтервал:
Закладка:








