Евгений Кунин - Логика случая. О природе и происхождении биологической эволюции

- Название:Логика случая. О природе и происхождении биологической эволюции
- Автор:
- Жанр:
- Издательство:Литагент «Центрполиграф»a8b439f2-3900-11e0-8c7e-ec5afce481d9
- Год:2014
- Город:Москва
- ISBN:978-5-227-04982-7
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Евгений Кунин - Логика случая. О природе и происхождении биологической эволюции краткое содержание
В этой амбициозной книге Евгений Кунин освещает переплетение случайного и закономерного, лежащих в основе самой сути жизни. В попытке достичь более глубокого понимания взаимного влияния случайности и необходимости, двигающих вперед биологическую эволюцию, Кунин сводит воедино новые данные и концепции, намечая при этом дорогу, ведущую за пределы синтетической теории эво люции. Он интерпретирует эволюцию как стохастический процесс, основанный на заранее непредвиденных обстоятельствах, ограниченный необходимостью поддержки клеточной организации и направляемый процессом адаптации. Для поддержки своих выводов он объединяет между собой множество концептуальных идей: сравнительную геномику, проливающую свет на предковые формы; новое понимание шаблонов, способов и непредсказуемости процесса эволюции; достижения в изучении экспрессии генов, распространенности белков и других фенотипических молекулярных характеристик; применение методов статистической физики для изучения генов и геномов и новый взгляд на вероятность самопроизвольного появления жизни, порождаемый современной космологией.
Логика случая демонстрирует, что то понимание эволюции, которое было выработано наукой XX века, является устаревшим и неполным, и обрисовывает фундаментально новый подход – вызывающий, иногда противоречивый, но всегда основанный на твердых научных знаниях.
Логика случая. О природе и происхождении биологической эволюции - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Эукариоты сильно различаются по характеру интронов: многие протисты и одноклеточные грибы содержат всего несколько интронов на весь геном, тогда как животные, растения и некоторые из простейших богаты интронами, так что кодирующие последовательности большинства их генов прерываются несколькими интронами (Jeffares et al., 2006). Замечательно, что позиции большой доли интронов консервативны у ортологичных генов разных организмов, включая растения и животных (Rogozin et al., 2003). Эволюционные реконструкции, принимающие во внимание консервативные и вариабельные позиции интронов, приводят к неожиданному выводу о том, что гены LECA были почти так же насыщены интронами, как и у современных млекопитающих, и значительная часть интронов LECA сохранилась по сей день в тех же позициях (см. рис. 7–8; Csuros et al., 2011). Этот вывод может казаться странным, но чем больше геномов становится доступно для анализа все более точными методами реконструкции, тем более убедительным он оказывается. Из этого наблюдения, формально подтвержденного результатами реконструкций, следует, что дальнейшая эволюция вела в первую очередь к утрате интронов, происходившей в большинстве ветвей эукариот, а немногие эпизоды взрывного увеличения их количества, по-видимому, были связаны с появлением новых крупных ветвей, таких как растения и животные (см. рис. 7–8). Резкое увеличение числа интронов у основания супергруппы Plantae могло быть обусловлено новой волной интронов группы II, перешедших от цианобактериального симбионта. Источник интронов у основания ветви животных остается загадочным и даже может свидетельствовать о роли скрытого эндосимбиоза в происхождении животных.
Реконструкция (см. рис. 7–8) была проведена с использованием методов Монте-Карло и марковских цепей (Csuros et al., 2011). Показана плотность интронов (число интронов на 1 Кб) для ныне существующих форм и предполагаемая плотность для ключевых предковых форм. Насыщенность черной штриховки приблизительно пропорциональна плотности интронов. Линия человека отмечена кружком. Показаны три супергруппы эукариот ( Chromalveolata, Unikonta и Plantae ) и основные группы внутри каждой из них, для которых известны полные последовательности геномов и соответствующие данные по локализации интронов [67].
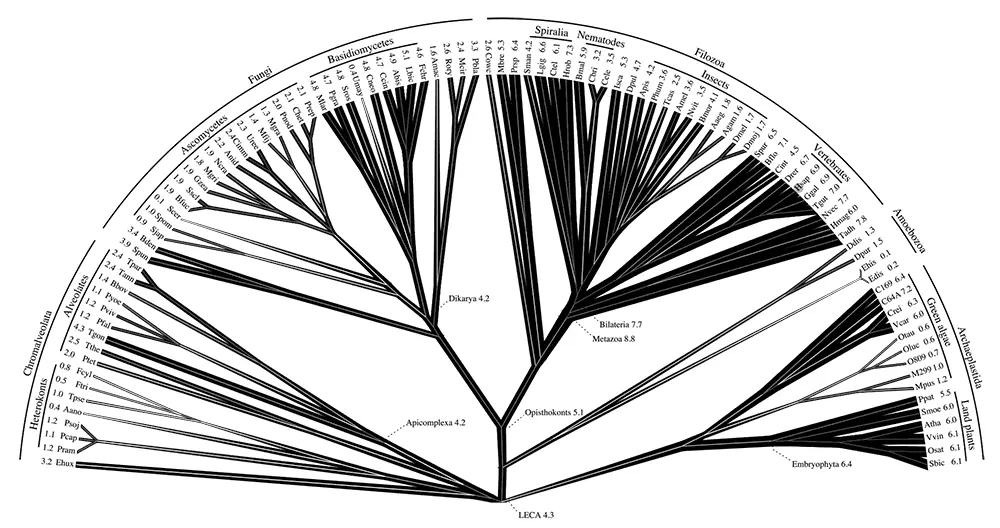
Рис. 7–8. Реконструкция приобретения и утраты интронов в течение эволюции эукариот и плотности интронов у предковых форм.
По: Csuros et al., 2011. Данная статья находится в свободном доступе под Атрибутивной лицензией Creative Commons .
Таким образом, LECA, по-видимому, аккумулировал интроны до плотности, близкой к наиболее богатым интронами современным геномам. Что же можно сказать о динамике интронов в течение стволовой фазы, между эукариогенезом и LECA? Простой расчет показывает, что, если инвазия интронов произошла «мгновенно», то протоэукариотический геном должен был большей частью (до 80 процентов) состоять из интронов, учитывая большие и единообразные размеры интронов группы II (около 2,5 Кб; Koonin, 2009b). Скорее всего, это чрезмерное упрощение. Процесс аккумуляции интронов, вероятно, был более постепенным и сопровождался уменьшением встроившихся интронов. Как бы то ни было, интроны, по всей видимости, сыграли ключевую роль в самом начале эволюции эукариот, согласно обсуждаемой модели эукариогенеза.
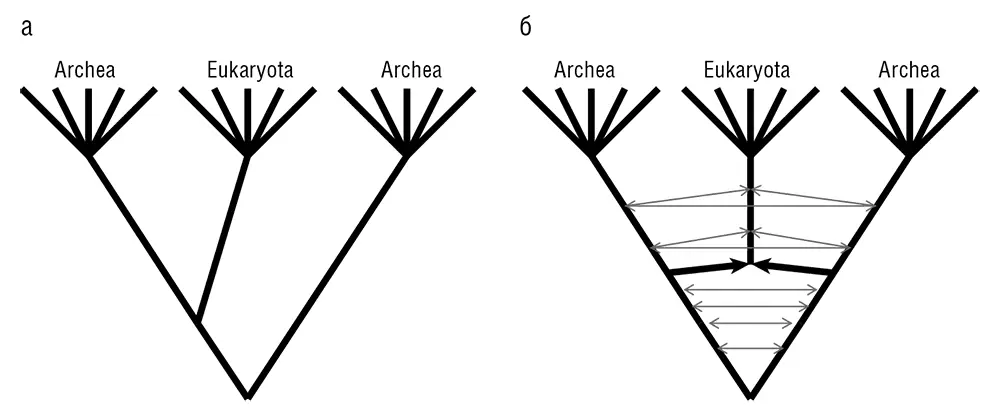
Рис. 7–9. Пересмотр трех доменов жизни: а – традиционное трехдоменное дерево Вёзе; б – циклический граф трех доменов согласно симбиогенетическому сценарию эукариогенеза и ГПГ между доменами.
Три домена жизни: за пределами дерева Вёзе
Симбиогенетический сценарий эукариогенеза ведет к пересмотру трехдоменного дерева жизни, разработанного Вёзе и его коллегами, даже если не учитывать последствия широкомасштабного ГПГ между прокариотами, что обсуждалось в главах 5 и 6. Дерево Вёзе подразумевает архезойный сценарий, а эндосимбиоз рассматривается как относительно позднее событие в истории эукариотического домена, которое не имеет отношения к трехдоменной классификации жизни (см. рис. 7–9 а ). Напротив, симбиогенетический сценарий постулирует, что захват первичного эндосимбионта буквально дал начало эукариотическому домену и, в процессе, внес большое число (возможно, большинство) генов в развивающийся эукариотический геном. Согласно последнему сценарию, слияние организмов из двух первичных доменов дало начало третьему домену; тогда конечная диаграмма не будет представлять собой дерево (см. рис 7–9 б ). Важное следствие, к которому мы вернемся в главе 11, состоит в том, что, размышляя о происхождении клеток, мы должны думать только о двух прокариотических доменах: археях и бактериях.
Краткий обзор главы
Среди трех доменов жизни эукариоты, без сомнений, обладают самой сложной, поразительно изощренной клеточной организацией, которая может даже навести некоторых на мысли о «неупрощаемой сложности» (Kurland et al., 2006), так как для большинства характерных функциональных систем эукариотических клеток мы не можем найти эволюционные промежуточные формы. Естественно, эукариогенез представляется одной из важнейших проблем эволюционной биологии, задачей, для решения которой «в лоб» мы гораздо лучше оснащены, чем для решения более фундаментальных задач происхождения клетки и, в конце концов, происхождения жизни (см. гл. 11 и 12). Сравнительная геномика на сегодняшний день не смогла ни разгадать загадку эукариогенеза, ни предложить исчерпывающую картину первичного расхождения главных эукариотических линий. Тем не менее филогенетический анализ во многих случаях внес ясность в вопросы, связанные с происхождением и самыми ранними этапами эволюции эукариот. Так, филогеномика прояснила эволюционные связи между царствами эукариот и привела к выявлению пяти или шести супергрупп. Родственные связи между супергруппами и корнем древа эукариот остаются чрезвычайно сложными для расшифровки, возможно по причине сжатого кладогенеза при первичном расхождении главных ветвей эукариот. Продолжающийся сбор данных по геномам различных ветвей жизни – отнюдь не игра в бирюльки; напротив, сравнительный анализ различных геномов продолжает приносить неожиданные биологические открытия, так что ожидать, безусловно, следует еще большего.
Ультраструктурные, функциональные и геномные данные согласованно свидетельствуют о том, что эукариоты являются археобактериальными химерами. Более того, гены явно бактериального происхождения существенно превышают по численности «архейные» гены. Более того, сравнительный анализ растущей коллекции доступных геномов архей все увереннее показывает, что многие ключевые клеточные системы эукариот существуют в примитивных формах уже у архей. Вариабельность этих систем между разными линиями архей, вместе с филогенезом консервативных белков, говорит о том, что архейный предок эукариот принадлежал к глубокой, возможно вымершей ветви архей с геномом высокой сложности и разработанными клеточными функциями. Недавнее открытие возможного прямого предка убиквитиновой системы в новом архейном геноме говорит о том, что мы можем в настоящее время недооценивать, насколько многие типичные функциональные системы эукариот могли быть представлены в течение эволюции Archaea . Это и другие подобные открытия внушают доверие к «комбинированному сценарию» происхождения эукариот, согласно которому эти заранее сформировавшиеся системы случайно сочетались в архейном хозяине первичного эндосимбионта. Напротив, системы внутренних мембран эукариот – особенно ядро, с его сложным комплексом ядерной поры – не обнаружены у архей и, по-видимому, были собраны, по крайней мере частично, из предковых бактериальных компонентов. Представляется важным, что эукариоты унаследовали развитые, сложно организованные системы от архей (естественно, за исключением митохондрий), тогда как многочисленные бактериальные молекулярные компоненты были в основном унаследованы по одному, a новые молекулярные машины возникли путем рекомбинации. Это различие, по-видимому, отражает асимметрию между хозяином и эндосимбионтом: несмотря на все драматические инновации, сопровождавшие эукариогенез, многие клеточные системы архейного хозяина сохранились и изменялись только эволюционным (плавным) путем, посредством дупликаций и приобретения дополнительных деталей.
Читать дальшеИнтервал:
Закладка:








