Евгений Кунин - Логика случая. О природе и происхождении биологической эволюции

- Название:Логика случая. О природе и происхождении биологической эволюции
- Автор:
- Жанр:
- Издательство:Литагент «Центрполиграф»a8b439f2-3900-11e0-8c7e-ec5afce481d9
- Год:2014
- Город:Москва
- ISBN:978-5-227-04982-7
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Евгений Кунин - Логика случая. О природе и происхождении биологической эволюции краткое содержание
В этой амбициозной книге Евгений Кунин освещает переплетение случайного и закономерного, лежащих в основе самой сути жизни. В попытке достичь более глубокого понимания взаимного влияния случайности и необходимости, двигающих вперед биологическую эволюцию, Кунин сводит воедино новые данные и концепции, намечая при этом дорогу, ведущую за пределы синтетической теории эво люции. Он интерпретирует эволюцию как стохастический процесс, основанный на заранее непредвиденных обстоятельствах, ограниченный необходимостью поддержки клеточной организации и направляемый процессом адаптации. Для поддержки своих выводов он объединяет между собой множество концептуальных идей: сравнительную геномику, проливающую свет на предковые формы; новое понимание шаблонов, способов и непредсказуемости процесса эволюции; достижения в изучении экспрессии генов, распространенности белков и других фенотипических молекулярных характеристик; применение методов статистической физики для изучения генов и геномов и новый взгляд на вероятность самопроизвольного появления жизни, порождаемый современной космологией.
Логика случая демонстрирует, что то понимание эволюции, которое было выработано наукой XX века, является устаревшим и неполным, и обрисовывает фундаментально новый подход – вызывающий, иногда противоречивый, но всегда основанный на твердых научных знаниях.
Логика случая. О природе и происхождении биологической эволюции - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
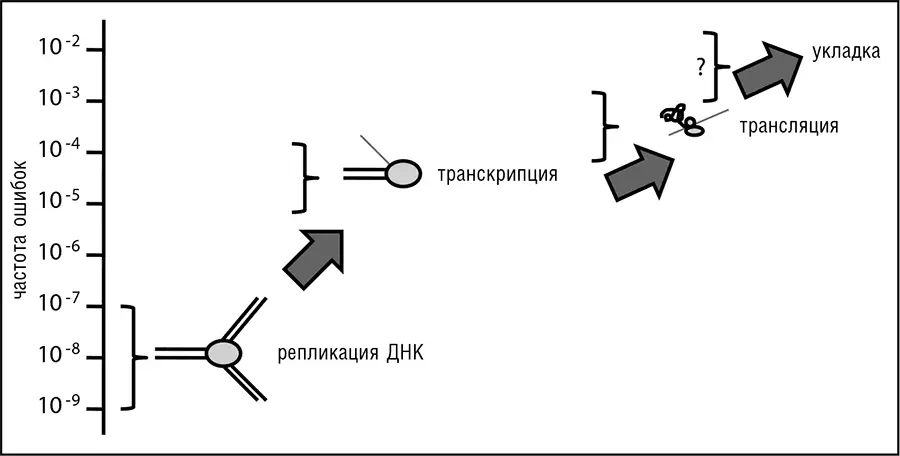
Рис. 9–4. Типичные частоты ошибок на различных стадиях передачи биологической информации.
Эксперименты с точными, но медленными рибосомными мутантами позволяют предположить, что компромисс между скоростью и точностью трансляции связан с механическими ограничениями трансляционной системы и с трудом может быть преодолен мутациями в ее компонентах. В результате для ограничения ошибок трансляции и их пагубных последствий эволюция, похоже, выбрала другие способы адаптаций.
Твердо установлено, что «высокостатусные» гены (те, что интенсивно экспрессируются и медленно эволюционируют) обладают более выраженным предпочтением триплетов по сравнению с «низкостатусными» генами. Оптимальные триплеты, которыми насыщены гены с высоким статусом, обеспечивают более низкую частоту ошибок, равно как и более высокую скорость трансляции, и, таким образом, частично избегают вышеупомянутого компромисса (Drummond and Wilke, 2009, 2008). Асимметрия триплетов между высокостатусными и низкостатусными генами может объясняться ценой селекции; в силу этого заметный отбор оптимальных кодонов может идти только в генах высокого статуса.
Основным вредным эффектом ошибок трансляции считается неправильная укладка белка (Drummond and Wilke, 2009, 2008), хотя ошибки включения аминокислот в каталитических сайтах, безусловно, могут стать дополнительным фактором. Как уже говорилось в главе 4, отбор на устойчивость к неправильной укладке является одним из главных аспектов эволюции белков – возможно, даже ее основной движущей силой. Менее ясно, какой источник неверной укладки наиболее важен – изначально неправильная последовательность или ошибки при ее трансляции. В любом случае, хотя укладка белка обычно не рассматривается в качестве процесса передачи информации, в действительности она им является. В самом деле, укладка предполагает поток информации, идущий от одномерной аминокислотной последовательности к трехмерной структуре белка .
Ровно то же самое относится к структурным РНК. Частоту неправильной укладки трудно определить экспериментально, и этого не было сделано для большого числа белков или РНК. Если тенденция, отражающая процент ошибок и показанная на рис. 9–4, – чем дальше от генома, тем менее точен этап передачи информации, – служит каким-либо индикатором, частота ошибок укладки должна быть даже выше частоты ошибок трансляции.
Такое предсказание также следует из здравого смысла, учитывая невероятную сложность процесса укладки и огромное количество ошибочных вариантов, доступных в принципе для укладываемого белка или РНК-молекулы (Bowman et al., 2011; Pande et al., 1998). Имея в виду высокую сложность пространства укладок, эпохальное открытие (сделанное первоначально Кристианом Анфинсеном и впоследствии подтвержденное многочисленными экспериментами), что белки способны самопроизвольно складываться в нативную конформацию, вызвало огромное удивление (Anfinsen, 1973).
Спустя почти 50 лет после открытия Анфинсена все еще остается предметом споров, глобальный или локальный минимум свободной энергии ищут спонтанно укладывающиеся белки. Но стало ясно, что лишь небольшие белки укладываются спонтанно; большинство белков нуждаются в специальных молекулярных устройствах, других белках, известных как шапероны, чтобы сформировать правильную структуру. Шапероны функционируют удивительным образом: их молекулы образуют «ячейку» (известную также как ячейка Анфинсена), которая изолирует укладываемый белок от цитоплазмы и частично разворачивает его, облегчая тем самым поиск нативной конформации (Ellis, 2003). Большинство шаперонов – синтезируемые в больших количествах, высококонсервативные, высокостатусные белки.
Первоначально некоторые из шаперонов были открыты как «белки теплового шока», то есть белки, которые резко усиливают свою активность при повышенной температуре (и, как было показано позже, при других стрессовых условиях) и противодействуют неправильной укладке других белков, которая усугубляется при стрессе (Vabulas et al., 2010). Хотя это явление менее детально исследовано, белковые шапероны также способствуют укладке молекул РНК (Russell, 2008; Woodson, 2010). В целом контроль над укладкой белков (и, вероятно, РНК) является, без сомнения, одной из основных функций во всех клетках.
Помимо устройств, подобных шаперонам, все клетки задействуют арсенал разнообразных молекулярных машин для контроля управляемого расщепления белков, в частности неправильно уложенных, и РНК. Как и молекулярные шапероны, эти машины – протеасомы, в случае белков, и экзосомы (деградосомы у бактерий) в случае РНК – повсеместно распространены во всех трех доменах жизни, присутствуют в изобилии в большинстве клеток и подвержены регулированию в условиях стресса (Hartung and Hopfner, 2009; Volker and Lupas, 2002; см. также гл. 7). Кроме того, эти машины, наряду с дополнительными вспомогательными системами регулируемого протеолиза, являются основными внутриклеточными потребителями энергии (АТФ). Бактерии дополнительно обладают высококонсервативными системами так называемой транс-трансляции, которые освобождают забуксовавшие рибосомы из аберрантных мРНК, на которых трансляция не в состоянии прекратиться должным образом, и предназначают такие мРНК и их белковые продукты (также аберрантные) к разрушению (Keiler, 2008).
Как мы подробно обсуждали в главе 7, эукариоты обладают важной стадией обработки информации, которая фактически не имеет эквивалента у прокариот: сплайсинг первичных транскриптов. Сопутствующая система контроля качества, по-видимому, эволюционировала одновременно с возникновением эукариот (см. гл. 7): механизм нонсенс-опосредованного распада (НОР), распознающий и уничтожающий аберрантные мРНК, которые содержат стопкодоны внутри экзонов помимо последнего, 3’-концевого экзона кодирующей последовательности (Behm-Ansmant et al., 2007; Stalder and Muhlemann, 2008).
Итак, контроль частоты ошибок и их влияния на биологические процессы передачи информации представляется одним из ключевых аспектов эволюции. По причинам, которые мы понимаем лишь частично (в лучшем случае), процент ошибок, по-видимому, не падает сильно ниже максимально допустимого значения: порога мутационного вырождения и соответствующего катастрофического порога фенотипических мутаций, который не изучен подробно, но предположительно существует. В случае частоты мутаций, простая неадаптивная теория популяционной генетики вполне способна объяснить наблюдаемые значения с достаточной степенью надежности (Lynch, 2010). Аналогичная аргументация была применена к фенотипическим мутациям (Burger et al., 2006), но в этом случае решение представляется менее очевидным.
Читать дальшеИнтервал:
Закладка:








