Даглас Хофштадтер - ГЕДЕЛЬ, ЭШЕР, БАХ: эта бесконечная гирлянда

- Название:ГЕДЕЛЬ, ЭШЕР, БАХ: эта бесконечная гирлянда
- Автор:
- Жанр:
- Издательство:Издательский Дом «Бахрах-М», 2001.
- Год:2001
- Город:Самара
- ISBN:ISBN 5-94648-001-4
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Даглас Хофштадтер - ГЕДЕЛЬ, ЭШЕР, БАХ: эта бесконечная гирлянда краткое содержание
Не часто приходится держать в руках книгу, которая открывает новые миры, в которой сочетаются глубина мысли и блестящая языковая игра; книгу, которой удалось совместить ничем на первый взгляд не связанные сложные области знания.
Выдающийся американский ученый изобретает остроумные диалоги, обращается к знаменитым парадоксам пространства и времени, находит параллели между картинами Эшера, музыкой Баха и такими разными дисциплинами, как физика, математика, логика, биология, нейрофизиология, психология и дзен-буддизм.
Автор размышляет над одной из величайших тайн современной науки: каким образом человеческое мышление пытается постичь самое себя. Хофштадтер приглашает в мир человеческого духа и «думающих» машин. Это путешествие тесно связано с классическими парадоксами, с революционными открытиями математика Курта Геделя, а также с возможностями языка, математических систем, компьютерных программ и предметного мира говорить о самих себе с помощью бесконечных отражений.
Начав читать эту книгу,вы попадете в волшебные миры, отправитесь в путешествие, изобилующее увлекательными приключениями, путешествие, после которого вы по-иному взглянете на мир и на самого себя.
Переведенная на 17 языков, книга потрясла мировое интеллектуальное сообщество и сразу стала бестселлером. Теперь и русский читатель получил доступ к одной из культовых книг XX века.
ГЕДЕЛЬ, ЭШЕР, БАХ: эта бесконечная гирлянда - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
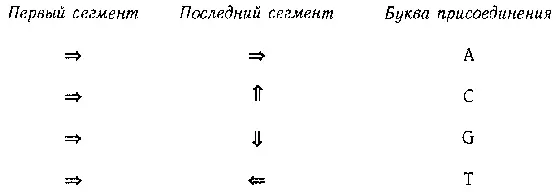
Рис. 89. Таблица «прикрепительных вкусов» типоэнзимов.
Таким образом, наш энзим предпочитает букву С. Иногда, складываясь, энзим пересекает сам себя — ничего страшного, просто представьте, что он проходит над или под собой. Обратите внимание, что все аминокислоты энзима играют роль в определении его третичной структуры.
Остается объяснить только одно. Почему в углу квадрата ААТипогенетического Кода нет никакой буквы? Дело в том, что дуплет ААдействует как знак препинания внутри цепочки, указывая на конец кода для данного энзима. Это означает, что в одной цепочке может быть закодировано несколько энзимов, если она содержит один или несколько дуплетов АА. Например, в цепочке:
CG GA ТА СТ АА AC CG А
закодировано два энзима:
кор — vsa — pmp — byk
и
raz — кор
ААразделяет цепочку на два «гена». Ген — это кусок цепочки, в котором закодирован один энзим . Заметьте, что не всякое ААявляется знаком препинания, например, CAAGделится на энзимы «sdp — str». ААначинается с четного подразделения и, таким образом, не составляет дуплета! Механизм, читающий цепочки и производящий закодированные в них энзимы, называется рибосомой . (Играя в типогенетику, мы проделываем работу рибосом.) Рибосомы не отвечают за третичную структуру энзимов, поскольку она полностью определена их первичной структурой. Процесс перевода всегда происходит от цепочек к энзимам , а не наоборот.
Теперь вы знаете правила типогенетики и можете поэкспериментировать с этой игрой. В частности, весьма интересно было бы попытаться получить самовоспроизводящуюся цепочку. Вот что это означало бы: дана некая цепочка; рибосома действует на нее, производя закодированные там энзимы. Затем эти энзимы вступают в контакт с первоначальной цепочкой и начинают с ней работать. Получается множество дочерних цепочек. Сами дочерние цепочки взаимодействуют с рибосомами, вследствие чего получаются новые энзимы, действующие на дочерние цепочки, и цикл продолжается. Наша надежда в том, что рано или поздно среди полученных цепочек мы найдем две копии первоначальной цепочки (на самом деле, одна из копий может оказаться самой первоначальной цепочкой.)
Схема типогенетических процессов представлена на следующей диаграмме.
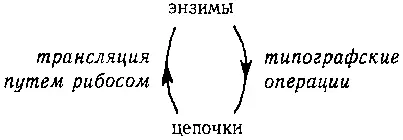
Рис. 90. «Центральная Догма типогенетики.» пример «Запутанной Иерархии».
На этой диаграмме показана Центральная Догма типогенетики . Из нее видно, как цепочки определяют энзимы (через Типогенетический Код) и как энзимы, в свою очередь, действуют на породившие их цепочки; в результате этого получаются новые цепочки. Таким образом, левая стрелка показывает, как старая информация подается наверх (ведь энзим является трансляцией цепочки и, следовательно, содержит ту же информацию, но в другой, активной форме). Правая стрелка, однако, не показывает движение информации вниз; вместо этого она указывает на то, как создается новая информация : передвижением символов в цепочке.
Энзим в типогенетике, подобно правилу вывода в формальной системе, механически переставляет символы в цепочке, не принимая во внимание никакого «значения», которое может заключаться в этих символах. Таким образом, здесь наблюдается интересное смешение уровней. С одной стороны, цепочки, поскольку на них воздействуют энзимы, играют роль данных (на это указывает правая стрелка); с другой стороны, они также диктуют, какие операции должны быть проделаны с данными и, таким образом, играют роль программ (на это указывает левая стрелка). Играющий в типогенетику действует как интерпретатор и процессор. «Круговая порука», связывающая «верхний» и «нижний» уровни в типогенетике, показывает, что нельзя сказать, что цепочки или энзимы находятся выше (или ниже) уровнем по сравнению друг с другом. С другой стороны. Центральная Догма системы MIU выглядит так:
правила вывода
↓ (типографские операции)
строчки
В системе MIU мы видим четкое разделение на уровни: правила вывода находятся уровнем выше, чем строчки. То же происходит в ТТЧ и во всех других формальных системах.
Однако мы видели, что и в ТТЧ, в некотором смысле, есть смешение уровней. Дело в том, что разделение на язык и метаязык оказывается не таким жестким высказывания о системе отражаются внутри самой системы. Если нарисовать диаграмму отношений между ТТЧ и ее метаязыком, у нас получится нечто, удивительно напоминающее Центральную Догму Молекулярной Биологии. На самом деле, наша цель — рассмотреть это сравнение как можно подробнее, для этого мы должны указать, в чем типогенетика совпадает с настоящей генетикой и в чем они различаются. Разумеется, настоящая генетика намного сложнее типогенетики, но «концептуальный скелет», который читатель получил, играя в типогенетику, будет очень полезен для путешествия по лабиринту действительной генетики.
Мы начнем с обсуждения отношений между «цепочками» и ДНК, что расшифровывается как «дезоксирибонуклеиновая кислота» ДНК большинства клеток находится в ядре — небольшом районе, защищенном мембраной. Гунтер Стент назвал ядро «тронным залом» клетки, в котором царит ДНК ДНК состоит из длинных цепей относительно простых молекул, называемых нуклеотидами .
Каждый нуклеотид состоит из трех частей: (1) фосфатная группа, лишенная одного атома кислорода (отсюда «дезокси» в названии кислоты), (2) сахар под названием «рибоза» и (3) основание . Именно основание отличает один нуклеотид от другого; таким образом, чтобы указать на нуклеотид, достаточно указать на его основание. В нуклеотидах есть четыре типа оснований:
A: аденин,
G: гуанин : пурины
C: цитозин,
T: тимин : пиримидины
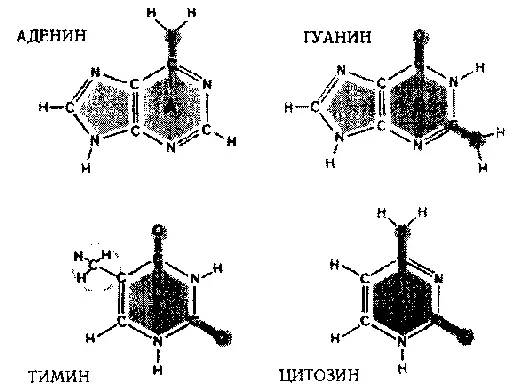
Рис. 91. Четыре основания, составляющих ДНК: Аденин, Гуанин, Цитозин, Тимин. (Hanawalt & Haynes. «The Chemical Basis of Life», стр. 142.)
(См. также рис. 91). Таким образом, цепочка ДНК состоит из множества нуклеотидов, следующих один за другим, как бусинки. Нуклеотид привязан к своим соседям сильной химической связью, которая называется ковалентной , «бусы» ДНК часто называются ее « ковалентным позвоночником ». ДНК обычно состоит из двух цепочек, чьи нуклеотиды спарены между собой (см. рис. 92).
Читать дальшеИнтервал:
Закладка:









