Александр Коган - Основы физиологии высшей нервной деятельности

- Название:Основы физиологии высшей нервной деятельности
- Автор:
- Жанр:
- Издательство:Высшая школа
- Год:1988
- Город:М.
- ISBN:5-06-001444-4
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Александр Коган - Основы физиологии высшей нервной деятельности краткое содержание
Учебник состоит из двух частей: общей физиологии высшей нервной деятельности, где изложены общие свойства условных рефлексов и нервных процессов, лежащих в их основе, и частной физиологии высшей нервной деятельности, где рассмотрены особенности условно-рефлекторного поведения основных классов животных, а также высшая нервная деятельность человека. 2-е издание (1-е — 1959 г.) значительно обновлено; особое внимание обращено на возможности использования высшей нервной деятельности для решения практических задач.
Максимальная ширина картинки в этой fb2-книге — 600 пикселей, но большинство картинок имеют размер не более 460 пикселей.
Основы физиологии высшей нервной деятельности - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Результаты некоторых экспериментов истолковывали как доказательство тормозящего действия стриарных структур на поведение животных. Однако дальнейшие исследования, в частности опыты с разрушением и стимуляцией хвостатых тел, и другие факты привели к заключению о наличии более сложных корково-подкорковых отношений.
Некоторые исследователи рассматривают факты об участии подкорковых структур в процессах высшей нервной деятельности как основание для того, чтобы считать их местом замыкания временных связей. Так возникло представление о «центрэнцефалической системе» как ведущей в поведении человека (У. Пенфилд, Г. Джаспер, 1958). В качестве доказательства замыкания временной связи в ретикулярной формации приводили наблюдения о том, что при выработке условного рефлекса первые изменения электрической активности мозга возникают именно в ретикулярной формации, а потом уже в коре больших полушарий. Но это свидетельствует лишь о вполне понятном раннем включении восходящей системы активации коры. Наконец, сильным аргументом в пользу подкорковой локализации замыкания считали возможность выработки условного, например зрительно-двигательного, рефлекса, несмотря на многократное рассечение коры на всю глубину, прерывающее все корковые пути между зрительной и двигательной областями. Однако этот экспериментальный факт не может служить доказательством, так как замыкание временной связи в коре имеет множественный характер и может происходить в любом ее участке между афферентными и эффекторными элементами. На рис. 9 жирными линиями показан путь условного зрительно-двигательного рефлекса при разрезах коры между зрительной и двигательной областями.
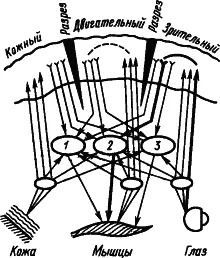
Рис. 9. Множественное замыкание временных связей в коре (показано пунктиром), которым не препятствуют ее разрезы (по А.Б. Когану):
1, 2, 3 — центральные механизмы оборонительных, пищевых и ориентировочных реакций соответственно; путь условного пищевого рефлекса на световой сигнал показан жирными линиями
Как показали многочисленные исследования, участие подкорковых структур в процессах высшей нервной деятельности не ограничивается регуляторной ролью ретикулярной формации среднего мозга и лимбических структур. Ведь уже на подкорковом уровне происходят анализ и синтез действующих раздражителей и оценка их биологического значения, в значительной мере определяющая характер образующихся связей с сигналом. Использование показателей формирования наиболее коротких путей, по которым сигнал достигает разных подкорковых структур мозга, выявило наиболее выраженное участие в процессах обучения задних отделов таламуса и поля CA 3гиппокампа. Роль гиппокампа в явлениях памяти подтверждается многими фактами. Наконец, нет оснований предполагать, что способность к примитивной замыкательной деятельности структур мозга, которая приобреталась в эволюции, когда они были ведущими, теперь у них полностью исчезла, когда эта функция перешла к новой коре.
Таким образом, корково-подкорковые отношения определяются регулированием функционального состояния коры активирующей системой — ретикулярной формацией среднего мозга и тормозной системой неспецифических ядер таламуса, а также возможным участием в образовании примитивных временных связей на нижнем уровне сложных иерархических механизмов высшей нервной деятельности.
Межполушарные отношения.Каким образом полушария головного мозга, являющиеся парным органом, участвуют в процессах образования условных связей? Ответ на этот вопрос был получен в опытах на животных, которым была сделана операция «расщепления» мозга путем перерезки мозолистого тела и передней комиссуры, а также продольного разделения перекреста зрительных нервов (рис. 10). После такой операции можно было вырабатывать разные условные рефлексы правого и левого полушарий, показывая разные фигуры правому или левому глазу. Если у оперированной таким образом обезьяны выработать условный рефлекс на световой стимул, подаваемый на один глаз, а затем подать его на другой глаз, то никакой реакции не последует. «Обучение» одного полушария оставляло другое «необученным». Однако при сохранении мозолистого тела и другое полушарие оказывается «обученным». Мозолистое тело осуществляет межполушарный перенос навыка.
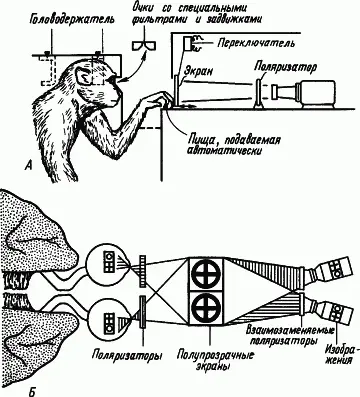
Рис. 10. Исследования процессов обучения у обезьян, подвергнутых операции «расщепления» мозга. А — устройство, направляющее в правый глаз одно изображение, а в левый — другое; Б — специальная оптика для проецирования зрительных образов в разные глаза (по Р. Сперри)
С помощью метода функционального выключения коры мозга у крыс воспроизводили на некоторое время условия «расщепленного» мозга. При этом временные связи могло образовать одно оставшееся активным полушарие. Этот рефлекс проявлялся и после прекращения действия распространяющейся депрессии. Он сохранился даже после инактивации полушария, которое было активным во время выработки этого рефлекса. Следовательно, «обученное» полушарие передало по волокнам мозолистого тела приобретенный навык «необученному». Однако этот рефлекс исчезал, если такую инактивацию производили до того, как полностью восстановится активность полушария, включенного во время выработки условного рефлекса. Таким образом, для переноса приобретенного навыка из одного полушария в другое необходимо, чтобы оба они были активны.
Дальнейшие исследования межполушарных отношений при образовании временных связей условных рефлексов показали, что во взаимодействии полушарий специфическую роль играют процессы торможения. Так, полушарие, противоположное стороне подкрепления, становится доминирующим. Оно вначале осуществляет формирование приобретенного навыка и его перенос в другое полушарие, а потом, притормаживая активность противоположного полушария и оказывая избирательно тормозное влияние на структуру временных связей, совершенствует условный рефлекс.
Таким образом, каждое полушарие, даже будучи изолированным от другого, способно к образованию временных связей. Однако в естественных условиях их парной работы сторона подкрепления определяет доминантное полушарие, которое формирует тонкую возбудительно-тормозную организацию условно-рефлекторного механизма приспособительного поведения.
Предположения о месте замыкания временных связей в больших полушариях головного мозга.Открыв условный рефлекс, И.П. Павлов сначала предположил, что временная связь является «вертикальной связью» между зрительной, слуховой или иными частями коры больших полушарий и подкорковыми центрами безусловных рефлексов, например пищевого — корково-подкорковая временная связь (рис. 11, А ). Однако многочисленные факты дальнейшей работы и результаты специальных опытов привели затем к выводу, что временная связь — это «горизонтальная связь» между очагами возбуждения, находящимися в пределах коры. Например, во время образования условного слюнного рефлекса на звук звонка замыкание происходит между клетками слухового анализатора и клетками, которые представляют в коре безусловный слюноотделительный рефлекс (рис. 11, Б ). Такие клетки назвали представителями безусловного рефлекса.
Читать дальшеИнтервал:
Закладка: