Александр Коган - Основы физиологии высшей нервной деятельности

- Название:Основы физиологии высшей нервной деятельности
- Автор:
- Жанр:
- Издательство:Высшая школа
- Год:1988
- Город:М.
- ISBN:5-06-001444-4
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Александр Коган - Основы физиологии высшей нервной деятельности краткое содержание
Учебник состоит из двух частей: общей физиологии высшей нервной деятельности, где изложены общие свойства условных рефлексов и нервных процессов, лежащих в их основе, и частной физиологии высшей нервной деятельности, где рассмотрены особенности условно-рефлекторного поведения основных классов животных, а также высшая нервная деятельность человека. 2-е издание (1-е — 1959 г.) значительно обновлено; особое внимание обращено на возможности использования высшей нервной деятельности для решения практических задач.
Максимальная ширина картинки в этой fb2-книге — 600 пикселей, но большинство картинок имеют размер не более 460 пикселей.
Основы физиологии высшей нервной деятельности - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Выработка условного торможения так же, как и выработка временных связей условного рефлекса, происходит в высших отделах нервной системы; у высших позвоночных — в коре и подкорковых структурах больших полушарий мозга.
Но высшие отделы мозга, в частности кора больших полушарий, имеют сложную структуру. Условный рефлекс образуется благодаря временной связи между анализаторами сигнального раздражения и механизмом подкрепляющей реакции; в каком же звене нейронных сетей, образующих пути осуществления условного рефлекса, вырабатывается условное торможение?
Результаты специальных исследований по изучению локализации условного торможения в корковых структурах мозга приводили ученых к разным взглядам (схематически представлены на рис. 31).
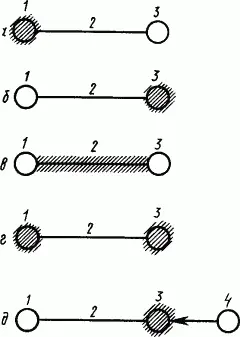
Рис. 31. Локализация внутреннего торможения (заштриховано) по представлениям различных авторов (по Л.Г. Воронину):
а — по Б.В. Бабкину, б — по И.Я. Перельцвейгу, в — по Э.А. Асратяну, г — по П.С. Купалову, д — по П.К. Анохину; 1 — корковая проекция условного раздражителя, 2 — временная связь, 3 — корковое представительство безусловного рефлекса, 4 — биологически отрицательная реакция
Первые исследования процесса угашения условного рефлекса, показавшие, что сигнальный стимул при определенных условиях перестает быть действующим, дали основание предполагать, что условное торможение развивается в зоне корковой проекции условного раздражителя (рис. 31, а ). Дальнейшие опыты со слюноотделительными рефлексами на подкрепление вливанием в рот раствора кислоты привели к заключению о задержке сигналов в «кислотном центре» и выводу, что условное торможение развивается в зоне корковой проекции безусловного раздражителя (рис. 31, б ). Так как на один и тот же сигнал можно было выработать два условных рефлекса: положительный и отрицательный, то, следовательно, зона проекции условного раздражителя не заторможена. В то же время на примере цепного двигательного рефлекса было показано, что зона проекции движении также остается назаторможенной. Это свидетельствовало в пользу мнения, что условное торможение развивается между зонами корковой проекции условного и безусловного раздражителей в структурах замыкания временной связи (рис. 31, в ).
Исследование взаимодействия условных слюнных рефлексов при их выработке на базе безусловных, вызываемых раздражением выведенных наружу лоскутков слизистой языка, привело к заключению, что условное торможение развивается в зоне корковой проекции как условного, так и безусловного раздражителей (рис. 31, г ). Наконец, исходя из того, что неподкрепление условного рефлекса вызывает «биологически отрицательную реакцию», которая может затормозить другую, одновременно протекающую деятельность, предположили, что условное торможение является внешним, которое «биологически отрицательная реакция» оказывает на зону корковой проекции безусловного раздражителя (рис. 31, д ).
Систематические исследования функционального состояния всех звеньев структуры условного рефлекса при его угашении привели к заключению, что ни в одном из них не возникал очаг торможения, который мог блокировать условное возбуждение, и оно в зоне проекции сигнального раздражителя становилось источником внутреннего торможения. Этому соответствует тот факт, что нейронные реакции коры мозга кошки при выработке дифференцировочного торможения оказались различными в зонах условного и безусловного раздражителей.
Разногласия по вопросу о том, в каком звене механизма условного рефлекса развивается условное торможение, крайне затрудняют суждение о его нейронной структуре. Однако можно с большой долей вероятности принять, что оно протекает в тех же нервных клетках, которые в свое время участвовали в образовании временной связи. Об этом свидетельствует, к примеру, такой факт, что при угасательном торможении корковый нейрон замедляет свой разряд с тем же латентным периодом, с которым он учащал разряд при положительном рефлексе. В опытах с клеточным аналогом временной связи выработка дифференцировочного торможения изменяла лишь ту часть импульсной реакции нейрона, которая составляла его условный ответ. В процессе развития внутреннего торможения существенную роль играет взаимодействие корковых элементов с подкорковыми, в частности со структурами неспецифической системы таламуса и ретикулярной формации среднего мозга.
Электрофизиологические показатели условного торможения.Когда условный рефлекс угашается, он просто прекращается. В этом является конечный результат деятельности торможения, но не сама деятельность. Некоторые стороны этой деятельности, скрытой от внешнего наблюдения, находят свое отражение в динамике ритмов основной электрической активности. Если у кошки с вживленными электродами вырабатывать на метроном 120 ударов/мин дифференцировку условного пищевого рефлекса подбегания к кормушке, образованного на метроном 60 ударов/мин, то произойдут характерные изменения в картине электрических потенциалов. При этом положительный условный раздражитель будет продолжать подавлять колебания основного ритма (6–8 колебаний/с) электрической активности и усиливать более быстрые колебания (30–40 колебаний/с). В то же время дифференцировочный раздражитель вскоре перестанет вызывать такие изменения основной электрической активности.
По мере дальнейшего развития дифференцировочного торможения неподкрепляемый сигнал при каждом своем применении будет усиливать колебания основного ритма и подавлять более быстрые колебания. При этом колебания основного ритма сами сильно замедлятся. Когда дифференцировка будет прочно выработана, отрицательный раздражитель — метроном с частотой 120 ударов/мин — не вызовет никакой двигательной реакции, но каждое его применение станет приводить к изменению электрической активности коры в сторону ее замедления. Часто при этом развивается дремотное состояние и возникают гигантские волны дельта-ритма. Однако медленные волны, наблюдаемые при угашении и выработке дифференцировок, рассматривают как необязательно связанные с развитием сна, а отражающие состояние неподвижности при бодрствовании, обозначая их «ритмами бодрствования — неподвижности». Отмечают, что при глубоком угашении условного рефлекса и выработке прочных дифференцировок медленные волны могут уменьшаться и электрическая активность возвращается к исходному состоянию.
Читать дальшеИнтервал:
Закладка: