Александр Коган - Основы физиологии высшей нервной деятельности

- Название:Основы физиологии высшей нервной деятельности
- Автор:
- Жанр:
- Издательство:Высшая школа
- Год:1988
- Город:М.
- ISBN:5-06-001444-4
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Александр Коган - Основы физиологии высшей нервной деятельности краткое содержание
Учебник состоит из двух частей: общей физиологии высшей нервной деятельности, где изложены общие свойства условных рефлексов и нервных процессов, лежащих в их основе, и частной физиологии высшей нервной деятельности, где рассмотрены особенности условно-рефлекторного поведения основных классов животных, а также высшая нервная деятельность человека. 2-е издание (1-е — 1959 г.) значительно обновлено; особое внимание обращено на возможности использования высшей нервной деятельности для решения практических задач.
Максимальная ширина картинки в этой fb2-книге — 600 пикселей, но большинство картинок имеют размер не более 460 пикселей.
Основы физиологии высшей нервной деятельности - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Все перечисленные особенности поведения корковых нейронов в разные стадии сна дают основание судить о нейрофизиологических механизмах его развития на нейронном уровне. Так, выключаемые из деятельности при засыпании нейроны, которые характеризуются полимодальностью, длинным латентным периодом и вариабельностью реакции, а также высокой частотой импульсации и повышенной возбудимостью, очевидно, имеют наиболее близкое отношение к осуществлению высших функций мозга. Можно предположить, что с их выключением связана потеря способности к активному восприятию окружающего и утрата сознания. Ограничение других функций мозга, возможно, обусловлено снижением возбудимости нейронов, замедляющих свои разряды, уменьшением информативности передаваемых сигналов, огрублением рабочих мозаик нейронной активности и перестройкой структуры импульсных потоков с преобладанием пачечного типа, характерного для восходящих таламокортикальных тормозных влияний.
Восстановление показателей нейронной активности в период ПС имело частичный характер. Можно предполагать, что это восстановление отражает растормаживание нейронных механизмов, имеющих отношение к явлениям, специфическим для ПС, как, например, быстрым движениям глаз или возникновениям сновидений. Однако для такого разграничения имеющихся сведений недостаточно, их могут дать дальнейшие исследования.
На рис. 45 приведена сводная схема расположения в структурах мозга систем, участвующих в регуляции уровня бодрствования и глубины сна. В соответствии с этой схемой последовательное включение нейрогуморальных механизмов, обусловливающих засыпание, смены стадий сна и пробуждение, можно представить по сигналам наступления темноты, прекращения деятельности и обстановки приготовления ко сну.
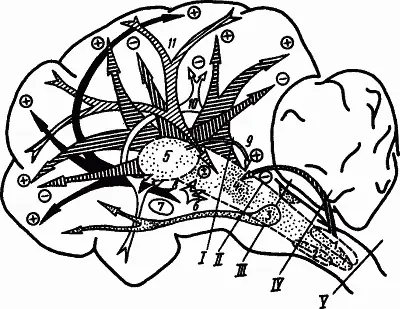
Рис. 45. Структуры мозга, участвующие в регуляции уровня бодрствования и глубины сна (по А.Н. Шеповальникову):
1 — синхронизирующая бульбарная система, 2 — «дополнительная» бульварная система, 3 — мостовой комплекс, обеспечивающий парадоксальный сон, 4 — активирующая ретикулярная формация ствола мозга, 5 — синхронизирующая таламическая система, 6 — активирующие влияния гипоталамуса на структуры ствола мозга, 7 — базальная синхронизирующая область, 8 — активирующие влияния гипоталамуса на структуры коры, 9 — облегчающее влияние высокочастотной стимуляции интраламинарных ядер таламуса на ретикулярную систему, 10 — влияния лимбической системы, способствующей сну, 11 — облегчающие и угнетающие влияния коры мозга на ретикулярную систему; I—V — уровни перерезок мозгового ствола при изучении нервных механизмов сна; знаком «+» обозначено активирующее действие, знаком «−» — угнетающее
Условное и безусловное торможение, которое развивается на фоне изменения активности гормонов и метаболизма, а также ограничение сенсорной информации и умеренное утомление обусловливают угнетение внешней деятельности путем ослабления влияния активирующих десинхронизирующих систем (рис. 45, 4, 6, 9, 11 ). Начинается засыпание.
Снижение тонуса активирующих систем помогает выявлению развивающейся к этому времени деятельности гипногенных синхронизирующих систем (рис. 45, 1, 2, 5, 7, 10 ). Гипоталамус оказывает трофотропное влияние на структуры ствола. В процесс вовлекается хвостатое ядро и утрачивается способность к активным движениям, блокируется поступление сенсорной информации. В нервных клетках ствола мозга, особенно в области синхронизирующей бульбарной системы, накапливается серотонин. Развивается медленноволновый сон.
Нарастающая на протяжении десятков минут медленноволнового сна деятельность гипногенных систем достигает степени, при которой (пока неясно, каким образом) происходит быстрый переход к активами центров дорсомедиальных отделов моста (рис. 45, 3 ), обусловливающих десинхронизацию в коре без участия ретикулярной формации среднего мозга. В соответствующих отделах мозга усиливается обмен веществ и повышается температура. Частичная активизация коры и ее связи с лимбической системой создают условия для появления сновидений. В структурах ствола мозга накапливается норадреналин и уменьшается серотонин. Возникает парадоксальный сон.
Активность нейрогуморальных механизмов парадоксального сна постепенно уменьшается и может на протяжении нескольких минут снизиться настолько, что усилившиеся за это время влияния гипногенных систем вызовут быструю обратную перестройку деятельности мозга в состояние медленноволнового сна. Однако это состояние, хотя и более продолжительное, чем состояние парадоксального сна, через некоторое время снова сменится им. Такое циклическое чередование стадий сна длится всю ночь, причем к утру продолжительность стадий парадоксального сна обычно возрастает.
С утренним рассветом связаны такие изменения активности гормонов и обмена веществ, которые в суточной периодике означают переход к активной внешней деятельности в светлое время. Под влиянием этих изменений, а также нарастающих потоков сенсорной информации и завершения восстановительных процессов в накануне работавших до утомления структурах мозга происходит снижение тонуса гипногенных систем, что благоприятствует резкому возрастанию деятельности активирующих систем, особенно десинхронизирующего действия ретикулярной формации среднего мозга. Наступает пробуждение.
В разные стадии сна по-разному протекают процессы высшей нервной деятельности. Так, выработанная на словесную инструкцию двигательная реакция сжатия руки в кулак сопровождалась изменениями ЭЭГ и ЭМГ, наиболее выраженными в ПС. Описаны нарушения памяти при устранении ПС. Однако приводятся доказательства того, что этим не исключается консолидация долгосрочной памяти при выработке реакций пассивного избегания.
В управлении сном принимает участие наряду с нервной гуморальная регуляция. Из аминокислотных остатков выделен дельта-пептид сна, вызывающий дельта-волны и характерный для МС (М. Монье, Г. Шененбергер, 1975). Из мозга животных, лишенных сна, выделен фактор S, увеличивающий продолжительность МС. В опытах на животных и в наблюдениях на людях показана роль серотонина и норадреналина в процессах сна. На основе этих данных предложен следующий механизм развития и прекращения сна: подавление симпатической активности приводит к поверхностному сну, расходование серотонина соответствует глубокому МС, расходование норадреналина — ПС, возбуждение симпатической активности вызывает пробуждение.
Читать дальшеИнтервал:
Закладка: