Александр Коган - Основы физиологии высшей нервной деятельности

- Название:Основы физиологии высшей нервной деятельности
- Автор:
- Жанр:
- Издательство:Высшая школа
- Год:1988
- Город:М.
- ISBN:5-06-001444-4
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Александр Коган - Основы физиологии высшей нервной деятельности краткое содержание
Учебник состоит из двух частей: общей физиологии высшей нервной деятельности, где изложены общие свойства условных рефлексов и нервных процессов, лежащих в их основе, и частной физиологии высшей нервной деятельности, где рассмотрены особенности условно-рефлекторного поведения основных классов животных, а также высшая нервная деятельность человека. 2-е издание (1-е — 1959 г.) значительно обновлено; особое внимание обращено на возможности использования высшей нервной деятельности для решения практических задач.
Максимальная ширина картинки в этой fb2-книге — 600 пикселей, но большинство картинок имеют размер не более 460 пикселей.
Основы физиологии высшей нервной деятельности - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Дафнии.Дафнии составляют основную массу планктона пресноводных водоемов. При недостатке корма или других неблагоприятных обстоятельствах они устремляются к свету, который сигнализирует о богатых пищей и более теплых поверхностных слоях воды. Однако при изменении условий жизни может измениться и сигнальное значение света.
Такое изменение, например, наблюдали в эксперименте при сочетании света с холодом, а тепла с темнотой.
Для этого сосуд с одной группой дафний при ярком освещении охлаждали до 2–4 °С и оставляли при низкой температуре в течение 4 ч. Затем его затемняли и нагревали до температуры 15–20 °С, оптимальной для дафний. У другой группы дафний вырабатывали обратные отношения, т.е. свет сочетался с теплом, а темнота с холодом. Соответствующие условные рефлексы быстро образовывались: дафнии первой группы шли в темноту, дафнии второй группы — к свету.
Изменить сигнальное значение света для дафний можно также, создавая иные условия питания.
Для этого помещали дафний в хорошо освещенный сосуд с чистой водой без корма. К вечеру их переносили в сосуд с водой, богатой инфузориями, закрывали светонепроницаемым колпаком и оставляли на всю ночь кормиться в темноте. Утром после ополаскивания дафний опять переносили в первый сосуд голодать весь день на свету и т.д. В результате у дафний выработался прочный условный пищевой рефлекс на темноту.
Ниже приведены данные по выработке условного пищевого рефлекса на темноту у дафний (по А.Б. Когану и А.П. Семеновых).
Число дней кормления в темноте и голодания на свету | Процент положительных реакций на темноту
0 | 0
7 | 32
14 | 57
21 | 89
Раки.Речные раки собираются к месту, где они несколько раз находили пищу. Этой способностью раков пользуются при их ловле, известна защитная реакция раков-отшельников, скрывающих свое мягкое брюшко в пустые раковины. Если ракам-отшельникам дать несколько раковин, отверстия которых забиты пробками, то после безуспешных попыток ввести брюшко в отверстия раковин раки прекращают это занятие. Такое торможение защитной реакции вырабатывается медленно. Пытаясь открыть раковины, раки упорно действуют клешнями в течение нескольких суток. Но когда торможение выработалось, оно оказывается весьма прочным. Даже спустя нескольких дней ракам предлагали те же раковины, но заклеенные тонкой бумагой вместо пробок, и бумага оставалась целой, отшельники даже не прикасались к объектам своей недавней энергичной деятельности. Однако если предложить им другие раковины, то раки немедленно их схватывают и начинают обследовать. Видимо, здесь происходит дифференцирование формы предмета.
Десятиногих раков испытывали в «лабиринте» (рис. 74, А ). Вначале число выходов в аквариум (1) и заходов в тупик (2) было примерно одинаковым, лишь постепенно раки стали правильно выбирать дорогу. Рис. 74, Б показывает «кривую обучения» раков. Даже после двух недель перерыва в работе условный рефлекс хорошо сохранился. Этот факт в соответствии с приведенными выше наблюдениями за поведением раков свидетельствует о значительной стойкости вырабатываемых временных связей.
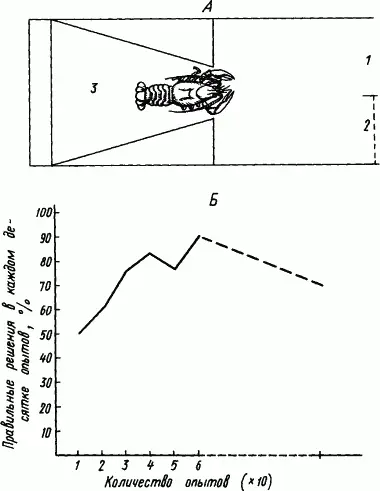
Рис. 74. Опыты «обучения» раков в простом лабиринте (по В.М. Боровскому). А — схема лабиринта; Б — кривая «обучения» рака в этом лабиринте:
1 — свободный вход в аквариум, 2 — стеклянная перегородка, закрывающая выход, 3 — выпускное отделение, прерывистая часть абсциссы означает 2-недельный перерыв
Когда речных раков стали кормить только в освещенной части аквариума, то у них за 4–12 сочетаний образовался и за 10–20 сочетаний укрепился условный пищевой рефлекс на освещение. Такой рефлекс удалось угасить за 6–17 применений светового сигнала без подкармливания, но на следующий день рефлекс восстанавливался.
Получены сведения о способности краба, где бы он ни находился, при испуге бежать по прямой линии к своей норе. Показано, что этот краб может использовать для ориентации сигналы поляризованного света. Однако скальный омар, покидая свое убежище для ночной кормежки, возвращается в него, используя, по-видимому, местные ориентиры. Вместе с тем некоторые наблюдения указывают на врожденную способность членистоногих к ориентации по неизвестным пока признакам. Так, когда особям береговой блохи, рожденным и выросшим в лаборатории, была предоставлена свобода передвижения, то они устремились в направлении берега, где обитали их родители.
Пауки.Условия охоты пауков сделали жужжание насекомых натуральными сигналами их охотничьей пищевой реакции. При этом паук живо реагирует на сотрясение паутины именно с той частотой, с которой муха бьет крыльями, пытаясь освободиться из плена. По-видимому, рассказы о «музыкальности» домового паука имеют известное основание. Некоторые тоны скрипки, близкие к мушиному жужжанию, привлекают паука так сильно, что он спускается как можно ближе к инструменту. Если подобрать камертон, имеющий сходную тональность, то, услышав его звучание, паук немедленно выглянет из своего убежища. Однако после нескольких таких применений камертона вырабатывается торможение реакции, не получающей пищевое подкрепление, звук теряет свое сигнальное значение.
Живущие в норах тарантулы подстерегают добычу у входа в нору. Они выскакивают и схватывают проходящих жуков, бабочек, стрекоз и других насекомых, но не реагируют на неподвижных.
Пищевой рефлекс паука очень силен и может быть использован для подавления защитного. Описывают, что даже свирепого калаб<...>ийского тарантула можно приучить к жизни в неволе в бумажной трубке, откуда он выходит и берет живых мух из рук человека.
Если предложить пауку-прыгуну муху, смазанную терпентином, то он ее не возьмет. Пищевой рефлекс будет заторможен. После того как пауку трижды давали таких мух, он переставал брать и обыкновенных мух, годных ему для еды. Это наблюдение, так же как и предыдущее, считают проявлением малой способности нервной системы паука к выработке дифференцировок. Однако когда паукам предлагали мух, пропитанных сахаром или хинином при звуках метронома 262 или 523 колебаний/с соответственно, то уже после 15 проб они переставали хватать мух при дифференцировочном звуке.
В пространственной ориентации пауков большую роль играет зрение. Так, паук при вибрации паутины от попавшего в нее насекомого бросается на него и, схватив, возвращается в свое убежище. При этом он ориентируется главным образом по направлению освещения. Если после того, как он схватил добычу, переместить источник света, то паук окажется дезориентированным и не сумеет сразу вернуться в убежище (рис. 75). По-видимому, обучение ориентации по направлению освещения происходит быстро, так как если источник света переместить даже незадолго до того, как паук бросится на добычу, то он уверенно возвращается в убежище.
Читать дальшеИнтервал:
Закладка: