Инал Акоев - Биофизика познает рак

- Название:Биофизика познает рак
- Автор:
- Жанр:
- Издательство:Наука
- Год:1988
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Инал Акоев - Биофизика познает рак краткое содержание
В книге на примере лейкоза человека рассмотрены в научно-популярном стиле и с позиции биофизики сложных процессов проблема рака, его причины, стадии развития и возможности предупреждения, ранней диагностики и лечения. Особое внимание уделено предраковым и предлейкозным состояниям организма, когда еще эффективна профилактика и еще не поздно изменить сложившиеся традиции и привычки в отношении своего поведения, характера питания и образа жизни, не способствующие укреплению противораковой устойчивости организма.
Книга предназначена для широкого круга читателей, интересующихся современными вопросами медицины и биологии.
Биофизика познает рак - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
В эритроцитарном направлении выявлено несколько промежуточных клеток (в том числе и бурстообразующая клетка) между общим предшественником миелопоэза и эритропоэтинчувствительными клетками. Последующие морфологически различаемые клетки относятся к проэритробласту, трем видам эритробластов, ретикулоциту и зрелому эритроциту. Обычно происходит 5—6 митозов в морфологически различимых клетках, и каждый эритробласт дает 30—60 эритроцитов. Эритробласты теряют способность к делению, как только в клетке накопится 27 нг гемоглобина. В случае замедления синтеза гемоглобина увеличивается число делений эритробластов. Из одной полипотентной стволовой клетки, начавшей дифференцировку в сторону миелопоэза, может образоваться около 1 млн эритропоэтинчувствительных клеток и около 0,1 млн клеток — предшественников гранулоцитов и макрофагов.
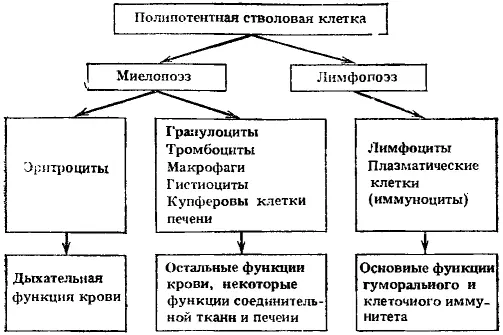
Рис. 3. Основные пути дифференцировки полипотентной стволовой клетки, клеточные формы и их главные функции
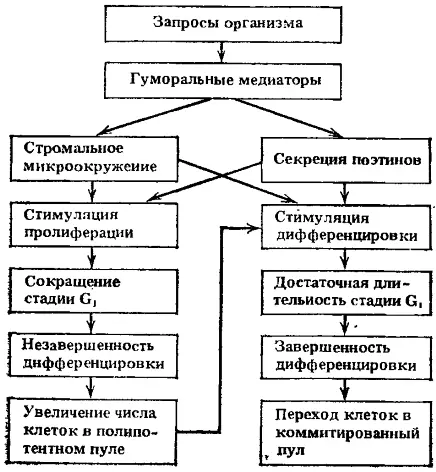
Рис. 4. Возможные пути регуляции пролиферации и дифференцировки полипотентных стволовых клеток в соответствии с запросами организма
Развитие гранулоцито-макрофагального направления миелопоэза проходит на стадии образования клеток, дающих колонии на полутвердых средах (KOE K), кластерообразующих клеток и предшественников гранулоцитов и моноцитов-макрофагов. Последующие клетки морфологически различимы. Миелобласт нейтрофильный далее дает клетки гранулоцитарного ряда (эезинофил), нейтрофильного ряда (нейтрофил) и базофильного ряда (базофил и тучная клетка).
Монобласт дает моноциты, макрофаги, гистиоциты, купферовы клетки печени и остеокласты. Клетки моноцитопоэтического ряда объединены в систему фагоцитирующих мононуклеаров как способные к фагоцитозу и осуществляющие в норме функции клеточного фагоцитоза. Ранее входившие вместе с указанными клетками в ретикуло-эндотелиальную систему ретикулярные клетки и эндотелиальные клетки исключены из системы фагоцитирующих мононуклеаров как являющиеся лишь факультативными фагоцитами, не имеющими рецепторов для иммуноглобулинов и не способными к иммунному фагоцитозу. Клетки же системы фагоцитирующих мононуклеаров в процессе дифференцировки приобретают рецепторы для иммуноглобулинов и комплемента и становятся способными к активному иммунному фагоцитозу.
Тромбопоэтинчувствительные клетки формируют морфологически различимые клетки: мегакариобласт, промегакариоцит и тромбоциты. Для предшественников характерны 4—5 эндомитозов.
Таким образом, по современным данным, клетки костного мозга, способные к делению, можно условно разделить на морфологически различимые клетки, как правило способные к очень ограниченному числу делений (4—8 делений), и на морфологически неразличимые клетки, выявляемые лишь по способности их давать колонии в селезенке облученных мышей или пролиферировать на полутвердых средах (и другими методами), способные к чрезвычайно активной пролиферации. Число делений, которые может совершить полипотентная стволовая клетка, на порядок больше, чем для морфологически различимых клеток. В норме их в костном мозге лишь около 0,5% от числа всех клеток. Однако достаточно сохраниться 0,1% стволовых клеток после сильного радиационного воздействия, чтобы обеспечить восстановление нормального пула стволовых клеток и резкое повышение дифференцировки клеток в последующих отделах.
При нормальном кроветворении абсолютное большинство стволовых клеток находится вне клеточного цикла (период G 0или длительный период G 1). Стабильное поддержание величины отдела стволовых клеток обеспечивается небольшой долей клеток, вступивших в митотический цикл. В случае уменьшения отдела стволовых клеток в пролиферацию может вовлекаться вся популяция сохранившихся стволовых клеток. При этом время удвоения популяции стволовых клеток оказывается очень коротким, порядка 15—20 ч. По данным ряда авторов, укорочение или удлинение стадии G 1стволовых клеток соответственно сопровождается пролиферацией (т. е. оставлением в пуле стволовых клеток) или дифференцировкой (т. е. уходом в последующие отделы).
Следовательно, основной вклад в восполнение дефицита клеточности костного мозга вносит пролиферация морфологически неразличимых клеток и потому исследование регуляции их численности и регуляции выбора направления дифференцировки является чрезвычайно важным. К сожалению, в этих вопросах еще очень много неясного.
Как решается для стволовых клеток первый выбор направления дифференцировки (в сторону лимфопоэза или миелопоэза), тоже неясно. По многочисленным данным, в опытах на собаках и обезьянах аллогенная кроветворная ткань вызывала появление огромного количества лимфоцитов, специфически иммунных против реципиента, при полной или частичной блокаде остального кроветворения. Следовательно, возможно переключение части стволовых клеток с эритроидного на лимфоидный путь дифференцировки. Характер дифференцировки кроветворной ткани в условиях трансплантации указывает на конкуренцию миелоидных и лимфоидных клеток за общий предшественник. У полицитемических радиохимер, у которых снижен эритропоэз, ускоряется восстановление лимфопоэза. Приведенные нами данные о существенной задержке пострадиационного восстановления числа лимфоцитов в крови человека в период, когда эритропоэз напряжен, также свидетельствуют о конкурентных отношениях ростков кроветворения. Об этом непосредственно говорит и проведенное нами изучение соотношений в морфологически различаемых клетках разных ростков костного мозга у облучавшихся собак.
Тем не менее некоторыми учеными предполагается, что направление дифференцировки стволовых клеток или закреплено генетически постоянно, или осуществляется стохастически, в обоих случаях — независимо от запросов организма. Возможным доказательством этого может служить одно и то же распределение маркерных клеток по разным росткам гемопоэза при изменении общих запросов организма.
В связи с изложенным, вероятно, более правильно считать существование конкурентных отношений между ростками кроветворения за преимущественную продукцию конечных зрелых форм клеток, а не примат дифференцировки полипотентной стволовой клетки в тот или иной росток гемопоэза. При этом влияние дальнодействующей для гемопоэза регуляции с целью реакций на запросы организма в экстремальных ситуациях может осуществляться не только через поэтинподобные регуляторы, но и через микроокружение в локальных участках гемопоэтических тканей, которое может зависеть также от запросов организма. Эритропоэтин для красного ростка и колониестимулирующий фактор для гранулоцитарного ростка стимулируют пролиферацию клеток и укорачивают средний период генерации клеток. При этом удлинение или укорочение стадии G 1клеточного цикла может определять уход клеток в пролиферацию или дифференцировку (рис. 4). В случае вступления клетки в деление сокращается генерационный цикл и клетка не успевает осуществить дифференцировку. Пролиферация прекращается, как только весь объем микроокружения окажется занятым стволовыми клетками.
Читать дальшеИнтервал:
Закладка:









