Инал Акоев - Биофизика познает рак

- Название:Биофизика познает рак
- Автор:
- Жанр:
- Издательство:Наука
- Год:1988
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Инал Акоев - Биофизика познает рак краткое содержание
В книге на примере лейкоза человека рассмотрены в научно-популярном стиле и с позиции биофизики сложных процессов проблема рака, его причины, стадии развития и возможности предупреждения, ранней диагностики и лечения. Особое внимание уделено предраковым и предлейкозным состояниям организма, когда еще эффективна профилактика и еще не поздно изменить сложившиеся традиции и привычки в отношении своего поведения, характера питания и образа жизни, не способствующие укреплению противораковой устойчивости организма.
Книга предназначена для широкого круга читателей, интересующихся современными вопросами медицины и биологии.
Биофизика познает рак - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Детально изучались возможные границы изменения этих факторов у млекопитающих. Здесь только укажем, что в условиях гомойотермного организма млекопитающих из двух указанных факторов, определяющих эффективность и полноту восстановления, в организме более оперативно и в течение более длительных периодов используется фактор времени. Так, блоковые задержки на стадиях клеточного цикла могут наступать немедленно вслед за повреждающим воздействием. В то же время клетки могут в определенных условиях уходить в период покоя G 0практически на неограниченное время.
Установлено, что в активно пролиферирующих реальных популяциях организма по мере усиления повреждения увеличивается продолжительность блоковой задержки. Однако она происходит лишь до определенного предела. Пока не достигнут предел продолжительности блоковой задержки, клетки, облученные в этой стадии, не имеют гибели ни интерфазного, ни митотического типа (в первом и последующих митозах). При отсутствии у клеток возможностей к дальнейшему увеличению продолжительности блоковой задержки в связи с усилением их повреждения (увеличение дозы облучения) они начинают гибнуть до митоза или во время последующих митозов.
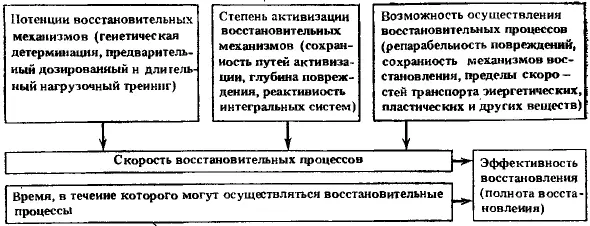
Рис. 10. Факторы, определяющие эффективность и полноту восстановления спонтанных и индуцированных внешним воздействием повреждений
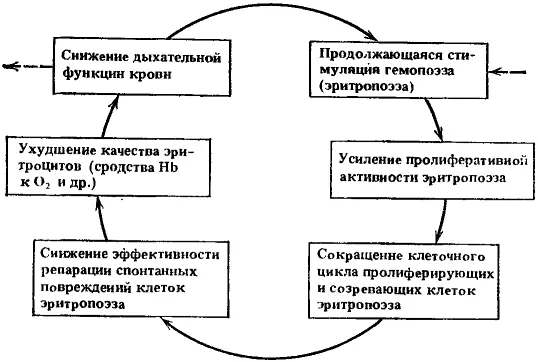
Рис. 11. Схема четвертого порочного потенциально патогенетического круга
Ведущие процессы: дальнейшая интенсификация эритропоэза и ухудшение качества эритроцитов
Следовательно, недостаток времени для осуществления восстановления клетки на данной стадии клеточного цикла от индуцированных повреждений заставляет клетку переходить в следующую стадию с сохранением части повреждения. Остаточные повреждения часто не мешают проходить даже очередной митоз, но они могут кумулироваться до уровня, несовместимого с жизнью клетки, и она погибает через один или несколько митозов.
При значительном усилении пролиферативной активности не всегда адекватный ответ ткани достигается только за счет вовлечения в делящийся пул клеток из покоящегося резерва. Происходит и сокращение продолжительности генеративного цикла клеток, в основном за счет периода G 1. При этом повышается чувствительность клеток к повреждающим воздействиям, например к радиации. Увеличивается и вероятность неполного восстановления спонтанных и индуцированных повреждений из-за недостатка времени на их репарацию. Клетки могут переходить из стадии в стадию и вступать в митоз с остаточными повреждениями, способными к кумуляции.
В этом мы видим одну из главных причин появления морфологических изменений (включений), совместимых с жизнью клетки и не имеющих определенного функционального значения. Вторая принципиально возможная причина появления таких изменений может заключаться в отставании поступления пластических и энергетических веществ от значительно ускоренных процессов роста и созревания клетки.
Выше были специально рассмотрены биохимические и биофизические изменения, сопровождающие ускорение процессов пролиферации и созревания клеток. Закономерные изменения, происходящие при значительном усилении пролиферативной активности, могут быть связаны с процессами генетической регуляции и могут носить внегеномный характер.
Наше внимание привлекли ошибки рибосомального синтеза белка, зависящие от изменения рибосомального окружения (ионный гомеостаз, pH и др.) и уменьшения времени на перебор кодон—антикодонных соотношений, происходящих, как предполагается, при сокращении продолжительности периода G 1. В этих случаях может включаться ошибочная аминокислота, близкая по своей кодовой специфичности «правильной» аминокислоте, информацию о которой несет матричная РНК. В таких случаях, несмотря на то что ни в ДНК, ни в матричной РНК, ни в рибосоме никаких изменений не произошло, происходит синтез белка с ошибками. Сократилось лишь время на один цикл работы рибосомы. Одна или несколько таких ошибок не могут существенно повлиять на активный центр фермента, но могут изменять антигенные свойства белка и модифицировать ряд физических характеристик белка и белоксодержащих структур (плазматических мембран и др.). Анализ всего пути следования информации от ДНК до функционирующего белка показал, что имеются и другие, но менее вероятные возможности внесения ошибок, влекущих за собой включение ошибочных аминокислот в пептидную цепь белка. Они во многом зависят от изменения окружения информационных молекул, от ионного гомеостаза, от pH и других локальных изменений, которые происходят при значительном усилении пролиферативной активности клеточной популяции и значительном ускорении клеточного деления.
При накоплении проходящих через митоз изменений могут уже ухудшаться качества белковой части гемоглобина, теряться сродство гемоглобина к кислороду, в связи с чем будет снижаться дыхательная функция крови. Это, в свою очередь, будет вновь усиливать стимуляцию эритропоэза.
Таким образом, сформировался новый порочный патогенетический круг, схема которого в упрощенном виде представлена на рис. 11.
Пятый порочный круг. Ведущие процессы: истощение эритропоэза и стимуляция лейкопоэза.Длительное функционирование четырех описанных выше порочных потенциально патогенетических кругов создает все возрастающую стимуляцию функций красной крови по изложенным уже механизмам межсистемных связей. В конце концов это может приводить к начинающемуся истощению эритропоэтической функции и явлениям стойкой анемии различной выраженности. Эта анемия некурабельна при применении гемостимуляторов.
Уменьшение числа эритроцитов и гемоглобина в периферической крови, снижение дыхательной функции крови приводят к мобилизации всех возможных резервов кроветворения для улучшения дыхательной функции. Продукция других ростков кроветворения снижена, начинает проявляться недостаточность лимфопоэза, гранулопоэза и тромбопоэза. В далеко зашедших случаях в крови может отмечаться снижение числа гранулоцитов, лимфоцитов, включая иммунокомпетентные клетки, и реже тромбоцитов. Картина изменений цитологического состава может быть весьма разнообразной. Чаще преобладает снижение зрелых клеточных форм одного из указанных ростков, но могут наблюдаться и сложные отношения, включая панцитопению. Проявления подобных состояний достаточно хорошо известны клиницистам.
Читать дальшеИнтервал:
Закладка:









