Николай Курчанов - Генетика человека с основами общей генетики. Учебное пособие

- Название:Генетика человека с основами общей генетики. Учебное пособие
- Автор:
- Жанр:
- Издательство:Литагент «СпецЛит»d5a9e1b1-0065-11e5-a17c-0025905a0812
- Год:2009
- Город:Санкт-Петербург
- ISBN:978-5-299-00411-3
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Николай Курчанов - Генетика человека с основами общей генетики. Учебное пособие краткое содержание
В пособии освещаются все разделы современной генетики, необходимые для понимания генетики человека и психогенетики. Показана методологическая роль генетики в современной биологии. Первые главы посвящены фундаментальным положениям общей генетики. В специальных разделах рассматриваются вопросы медицинской генетики, генной инженерии, генетики поведения, эволюции, психогенетики.
Второе издание книги значительно переработано автором с учетом новой информации, опубликованной за последние три года.
Пособие предназначено для студентов биологических, педагогических, психологических и социологических факультетов. Представляет интерес для научных работников всех специальностей, занимающихся вопросами, связанными с изучением биологической природы человека.
2-е издание, переработанное и дополненное.
Генетика человека с основами общей генетики. Учебное пособие - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Все рибосомы состоят из двух субъединиц – малой и большой (рис. 6.4). Размер рибосом и их субъединиц выражается скоростью седиментации частиц в растворе ( S – константа Сведберга ). Рибосомы прокариот характеризуются значениями 70S (30S + 50S), эукариот – 80S (40S + 60S). Рибосомы хлоропластов и митохондрий похожи на рибосомы прокариот, но отличаются значительной вариабельностью по размерам. Рибосомы содержат два участка – А (аминоацильный) и Р (пептидильный), являющимися основными каталитическими центрами. Помимо них, имеются и другие центры связывания ферментов. Специфичность участков определяется сочетанием соответствующих областей обеих субъединиц. При диссоциации субъединиц их специфичность теряется.
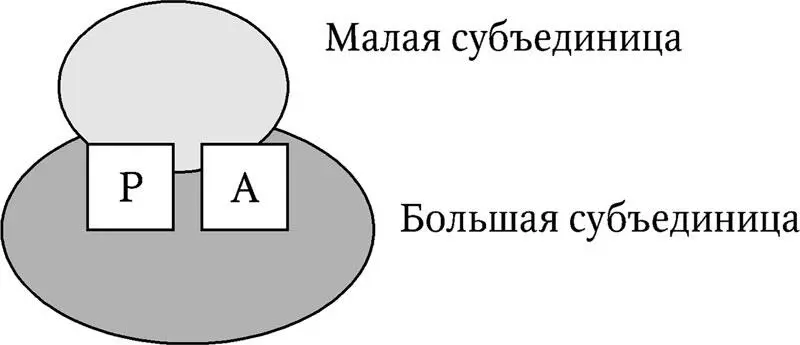
Рис. 6.4. Структура рибосомы
Как уже говорилось выше, рибосомы эукариот имеют в своем составе 4 разновидности р-РНК. Три из них образуются из единого предшественника (45S-РНК), синтез которого происходит в специализированной ядерной структуре – ядрышке– при помощи РНК-полимеразы-1. В ядрышках эукариот концентрируются петли хромосом, содержащие гены р-РНК. Эти гены обычно имеют много копий. Так, у человека 200 копий генов р-РНК располагаются на концах 5 пар хромосом (т. е. они имеются на 10 хромосомах из 46), поэтому сразу после митоза можно видеть 10 маленьких ядрышек, которые быстро сливаются в одно большое. Синтез 4-й р-РНК (5S-РНК) происходит вне ядрышка (у человека на 1-й хромосоме).
После поступления м-РНК на рибосомы и начинается процесс трансляции, в котором выделяют 4 стадии:
1. Стадия активации аминокислот. Активация свободных аминокислот осуществляется при помощи особых ферментов (аминоацил-т-РНК-синтетаз) в присутствии АТФ. Для каждой аминокислоты существуют свои ферменты и свои т-РНК. Активированная аминокислота присоединяется к своей т-РНК с образованием комплекса аминоацил-т-РНК (аа-т-РНК). Только активированные аминокислоты способны образовывать пептидные связи и формировать полипептидные цепочки.
2. Инициация. Начинается с присоединения лидирующего 5'-конца и-РНК с малой субъединицей диссоциированной рибосомы. Соединение происходит так, что стартовый кодон (обычно АУГ) оказывается в «недостроенном» Р-участке. Комплекс аа-т-РНК с помощью антикодона т-РНК присоединяется к стартовому кодону и-РНК. Имеются многочисленные (особенно у эукариот) белки – факторы инициации.
У прокариот стартовый кодон кодирует N-формилметионин, а у эукариот – N-метионин. В дальнейшем эти аминокислоты вырезаются ферментами и не входят в состав белка. После образования инициирующего комплекса происходит объединение субъединиц и «достраивание» Р– и А-участков рибосом.
3. Элонгация. Начинается с присоединения в А-участке и-РНК второго комплекса аа-т-РНК с антикодоном, комплементарным следующему кодону и-РНК. На рибосоме оказываются две аминокислоты, между которыми возникает пептидная связь. После соединения аминокислот первая т-РНК освобождается от аминокислоты и покидает рибосому. Рибосома перемещается вдоль нити и-РНК на один триплет (в направлении 5' → 3'). Вторая аа-т-РНК перемещается в Р-участок, освобождая А-участок, который занимает следующая аа-т-РНК. Таким же образом присоединяются 4-я, 5-я и т. д. аминокислоты, принесенные своими т-РНК.
4. Терминация. Завершение синтеза полипептидной цепочки. Наступает тогда, когда рибосома дойдет до одного из терминирующих кодонов. Имеются особые белки (факторы терминации), которые узнают эти участки.
На одной молекуле и-РНК может располагаться несколько рибосом (такое образование называется полисома), что позволяет осуществлять синтез нескольких полипептидных цепей одновременно.
Процесс биосинтеза белка проходит с участием большего количества специфических биохимических взаимодействий. Он представляет собой фундаментальный процесс природы. Несмотря на чрезвычайную сложность (особенно в клетках эукариот), синтез одной молекулы белка длится всего 3–4 секунды.
По аналогии с геномом, в последнее время получает распространение термин «протеом» как совокупность функциональных белков клетки. Необходимо заметить, что если гены во всех клетках организма одинаковы, то наборы белков весьма разнообразны, меняясь в разных типах клеток по ходу онтогенеза.
6.5. Перенос генетической информации в природе
Концепция генетического кода имела принципиальное значение для генетики, поскольку постулировала существование в природе определенной системы передачи информации. Пути переноса информации легли в основу представлений, которые Ф. Крик назвал центральной догмой молекулярной биологии.Основной порядок переписывания генетической информации в живой природе стал выражаться формулой:
ДНК → РНК → белок.
Такой поток информации реализуется в подавляющем большинстве живых систем. Единство генетического кода и общность потока генетической информации в природе показали, что эти явления произошли на самом раннем этапе эволюции жизни. Однако оба этих фундаментальных процесса имеют свои исключения. Вариации генетического кода мы рассмотрели выше. У некоторых вирусов были обнаружены особые варианты переноса информации – от РНК к РНК и от РНК к ДНК.
Огромное теоретическое значение имело открытие обратной транскрипции,вызвавшее широкий резонанс в научных кругах. Это открытие резко меняло устоявшиеся в генетике представления. При обратной транскрипции на РНК-матрице синтезируется двунитиевая структура ДНК (сначала одна нить, затем – другая). Этот процесс катализируется ферментом обратной транскриптазой (или РНК-зависимой-ДНК-полимеразой) . Обратная транскриптаза была открыта в 1970 г. (Baltimore D., 1970; Temin H., Mizutani S., 1970) при исследовании так называемых ретровирусов. Синтезированная ими молекула ДНК встраивается в хромосому клетки-хозяина и при делении передается дочерним клеткам. В такой форме геном вируса может существовать долгое время, переходя от родителей к потомкам. Скрытая интегрированная форма вируса получила название провирус.При активации провируса происходит воспроизведение РНК ретровируса. Эта РНК может транслироваться, синтезируя вирусные белки (в том числе и обратную транскриптазу ), и образовывать новые вирусные частицы, покрытые оболочкой.
Явление обратной транскрипции оказалось отнюдь не экзотикой. В настоящее время ретровирусам отводится определенная роль в эволюционном процессе как переносчикам генетической информации между организмами разных систематических групп. При образовании новых вирусных частиц активированного провируса может захватываться часть генетического материала клетки-хозяина и передаваться другим организмам, минуя межвидовые барьеры. Эволюционная роль ретровирусов будет рассмотрена ниже.
Читать дальшеИнтервал:
Закладка:









