Феликс Филатов - КЛЕЙМО СОЗДАТЕЛЯ. Гипотеза происхождения жизни на Земле.

- Название:КЛЕЙМО СОЗДАТЕЛЯ. Гипотеза происхождения жизни на Земле.
- Автор:
- Жанр:
- Издательство:ОАО ордена Знак почета Смоленская областная типография им. В.И.Смирнова
- Год:2011
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Феликс Филатов - КЛЕЙМО СОЗДАТЕЛЯ. Гипотеза происхождения жизни на Земле. краткое содержание
КЛЕЙМО СОЗДАТЕЛЯ. Гипотеза происхождения жизни на Земле. - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Этот рисунок иллюстрирует румеровское преобразование, переводящее дублеты одного октета в другой. Третье основание кодона неявно присутствует здесь в составе октета II, продукты которого организованы в две строки: верхнюю кодируют триплеты с третьим пиримидином, нижнюю — с третьим пурином.
Идеи Юрия Румера были продолжены и развиты работами Владимира Щербака. Два румеровских октета Щербак преобразовал таким образом, чтобы выделить в них группы вырожденности, пронумеровав их справа налево, а продукты кодирования (аминокислоты) он упорядочил в каждой группе по нарастанию молекулярной массы слева направо; триплеты, соответствующие продуктам кодирования, он записал по вертикали сверху вниз. Тогда первые, вторые и третьи основания кодонов образовывали три строки в каждом кодоне. Вот что у него получилось (цифры под третьими основаниями — характеристики кодируемого продукта):
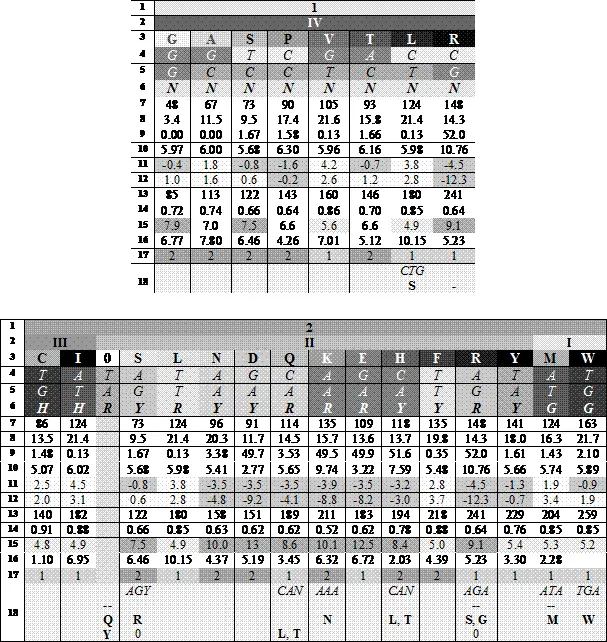
— номер октета
— номер группы вырожденности
— продукт кодирования ( аминокислотаили терминирующий сигнал 0)
— 1-е основание кодона
— 2-е основание кодона
— 3-е основание кодона
— Bulkiness(мера формы или объёма или «громоздкости» боковой цепи)
— Объем(рассчитанный по Ван-дер-Ваальсовым радиусам)
— Полярность(сила электрического поля вокруг молекулы)
— Изоэлектрическая точка
— Гидрофобность1; гидрофильные аминокислоты — выделены темносерым и отрицательными значениями
— Гидрофобность2 (другие данные)
— Поверхность, доступная для водыв развернутом пептиде
— Доступная воде поверхность, теряемая при свертываниипептида
— Polarrequirement , PR(эмпирические данные по хроматографии водных растворов). Темный >7.5; светлый <5.6
— Частота встречаемостиаминокислот в белках современных организмов
— ars -класс
— неканоническое кодирование(— нет продукта)
В двух прямоугольных блоках — два румеровских октета (строка 1), упорядоченные по нарастанию номера, каждый из которых разделен на группы вырожденности, помеченные соответствующими римскими цифрами (строка 2) и упорядоченные по убыванию номера. Строка 3 — продукты кодирования, упорядоченные по нарастанию масс в «своих» группах вырожденности (что — как и в первых двух случаях — подчеркивается градиентом насыщенности серого цвета). Символы оснований кодирующих триплетов расположены вертикально — сверху вниз, от 5` до 3` - для удобства сравнения кодонов по первым, вторым и третьим буквам соответственно. Четное содержание продуктов кодирования в октетах позволяет провести посередине вертикаль, которая оказывается осью симметрии по отмеченным ниже характеристикам. R — пурины ( A , G ), Y — пиримидины ( С , Т ), N — любое из четырех азотистых оснований. 0 — стоп-кодон. Строки 7-15 —физико-химические свойства аминокислот.
Эта несложная организация приводит к поразительно красивой общей картине: в октете 1первые основания триплетов (первая строка) оказываются взаимно комплементарными относительно упомянутой вертикали; вторые основания — зеркально симметричны по пуринам и пиримидинам. Симметрия третьих оснований — это симметрия их монотонного ряда. Симметрии подчеркиваются оттенками серого (интенсивность которого нарастает с увеличением молекулярной массы продукта кодирования в составе группы вырожденности).
Симметрия кодирующих триплетов и продуктов кодирования, относящихся к октету 2, в этой таблице немногосложнее, но также вполне наглядна. Она требует двух предварительных пояснений. Первое — это позиция цистеина, С. Универсальный код предписывает дублету TG кодирование цистеина, если третьим основанием кодона является пиримидин Y , а кодирование триптофана, если третья буква кодона — G . Аденин в третьей позиции образует пунктуационный знак — стоп-кодон TGA . То обстоятельство, что при этом нарушается симметрия, ставит определенную проблему, которую Владимир Щербак разрешил, обнаружив, что обозначенная в таблице позиция цистеина принципиально не противоречит Природе, поскольку существуют одноклеточные микро-организмы ( Euplotida , реснитчатые, инфузории), генетический код которых отличается от универсального как раз по кодированию цистеина: триплет TGA у них транслируется как Си не имеет функции « стоп ». Второе пояснение относится именно к функции « стоп », которая рассматривается в таблице как «законный» продукт кодирования, не имеющий массы.
Итак, упорядочивание продуктов кодирования октета 2по нарастанию молекулярной массы так же, как и в случае октета 1, приводит к симметриям первых, вторых и третьих оснований соответствующих кодонов. При этом основной особенностью этого представления является зеркальная симметрия пар кодирующих дублетов пяти краевых позиций и симметрия со сдвигом трех пар внутренних, кодирующих триплетов. Так же, как и в октете 1, осью этой симметрии является вертикаль, которая делит строки точно посередине. Симметрию третьих оснований нечетных групп вырожденности ( H | G ) Владимир Щербак рассматривал в данном случае как вариант симметрии Y | R .
Как мы упоминали, симметрияописанного представления генетического кода имеет место не только по молекулярной массе аминокислот, но и по другим их параметрам (строки 11-15).Замена в третьей позиции пары симметричных кодонов пурина пиримидином — и наоборот — в определенной мере сохраняет, например, гидрофобность кодируемых продуктов, хотя размер аминокислоты при этом, конечно, основательно меняется. Однако, ни гидрофобность, ни еще одна характеристика — PR (строка 15) — которую Карл Вёзе и его группа описали, как основу регулярности генетического кода и соответствия кодонов и продуктов кодирования, не могут сравниться по строгости симметрий с молекулярной массой аминокислот, что хорошо демонстрирует приведенная таблица Владимира Щербака. Неканонические ключи кодирования (строка 17) тоже выглядят в этих таблицах довольно беспорядочными и случайными отклонениями.
За весь этот рисунок кооперативной симметрии генетического кода , на основе молекулярных масс его компонентов, за ее красоту и гармонию, не имеющую к тому же сколько-нибудь внятного физического, химического или молекулярно-биологического обоснования, мой друг назвал описанную таблицу каллиграммой ( красивой записью ). Термин этот принадлежит Гийому Апполинеру, который — в экспериментальном порядке — попытался организовать некоторые свои тексты так, чтобы продемонстрировать их симметрию. Их он и называл calligrammes .
Читать дальшеИнтервал:
Закладка:





