Владимир Бердников - Эволюция и прогресс

- Название:Эволюция и прогресс
- Автор:
- Жанр:
- Издательство:Наука
- Год:1991
- Город:Новосибирск
- ISBN:5-02-030062-4
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Владимир Бердников - Эволюция и прогресс краткое содержание
Автор вводит читателя в круг наиболее интригующих вопросов эволюционной биологии. До сих пор эволюционный прогресс остается предметом бурных, даже ожесточенных споров. По существу, всех биологов можно разделить на сторонников и противников идеи этой формы прогресса. Эволюцию живых организмов обычно связывают с ростом их сложности и степени совершенства, однако до сих пор нет строгих критериев этой оценки. Главная мысль, развиваемая автором, состоит в том, что основные атрибуты прогресса — усложнение строения и повышение уровня надклеточной организации — являются лишь следствием постоянно идущего отбора на повышение эволюционной пластичности видов.
Книга предназначена для биологов широкого профиля, а также всех интересующихся вопросами эволюции живых существ.
Эволюция и прогресс - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
ΔX / X ≈ ΔlnX. (4.15)
Распределение, которое становится нормальным после логарифмирования значения случайной величины, называется логарифмически-нормальным. Его характерной особенностью является линейная связь между сигмой и средним значением. Вообще говоря, логарифмически-нормальное распределение совсем не симметрично и обладает длинным «хвостом», плавно спускающимся в сторону увеличения признака. Однако так оно выглядит при большом размахе изменчивости признака в арифметической шкале (например, в 10 раз). Если же такой диапазон значительно скромнее (скажем, только в 1,5 раза), то распределение по признаку в обеих шкалах выглядит очень сходным, и даже в арифметической шкале не слишком отличается от нормального. Поэтому при анализе особей одной популяции, когда размах изменчивости невелик, логарифмически-нормальные распределения выглядят как нормальные.
Опыт биометрии свидетельствует, что наследование количественных признаков, измеренных с помощью логарифмической шкалы, часто удовлетворяет аддитивной модели. Это означает, что межлинейное различие по величине логарифма признака можно трактовать как сумму эффектов аллельных замещений по ряду локусов. Заметим, что логарифм числа представим в виде суммы близких по величине слагаемых, когда само это число является произведением близких по значению сомножителей. Выходит, что замещение слабого аллеля на сильный в локусе, ответственном за развитие количественного признака, ведет к увеличению его генотипического значения в какое-то число раз. Причем это число не слишком различается при аллельных замещениях в разных локусах.
Ответ на искусственный отбор
Сначала рассмотрим, что кроется под таким образным понятием, как давление искусственного отбора. Проще всего под ним понимать долю отбракованных особей ( I ), обычно измеряемую в процентах:
I = (N 0/ N) 100 %, (4.16)
где N — численность популяции до отбора; N 0 — число отбракованных особей. Недостаток формулы (4.16) состоит в отсутствии какой-либо информации о признаке, по которому ведется отбор. Эту сторону работы селекционера передает так называемый селекционный дифференциал (S), который определяется как разница средних значений признака в исходной популяции (до браковки) ( М ) и у «счастливцев», отобранных для развода (М с), т. е.
S = M c— M. (4.16)
Аналогично можно ввести представление об ответе популяции на отбор (R) как о сдвиге среднего значения популяции за одно поколение отбора:
R = M' c— M , (4.18)
где M' c — среднее значение признака у потомков «счастливцев».
Большой экспериментальный материал свидетельствует, что ответ на отбор прямо пропорционален селекционному дифференциалу. Следовательно, чем сильнее давление отбора, тем больше величина ответа на него. Величина этого ответа — внутреннее (генетически обусловленное) свойство популяции. Его количественной мерой является наследуемость — коэффициент пропорциональности ( h 2 ), связывающий ответ на отбор с селекционным дифференциалом. Итак, при ответе конкретной популяции на отбор по конкретному количественному признаку выполняется равенство
R = h 2S. (4.19)
Наследуемость разных признаков у особей одной популяции может существенно различаться. Кроме того, может различаться и наследуемость одного и того же признака в разных популяциях одного вида. Что же лежит в основе этого явления?
Пусть мы имеем дело с популяцией, состоящей из генетически идентичных особей. Многократно показано, что при любом давлении отбора среднее значение любого признака в следующем поколении практически не изменяется ( h 2 = 0). Впервые это было продемонстрировано в знаменитом эксперименте В. Иогансена на чистых линиях фасоли. Иными словами, наследуемость равна нулю, если равна нулю генетическая компонента изменчивости.
Представим на момент, что мы в состоянии по фенотипу особи опознать ее генотип. Если к тому же эти особи размножаются бесполым путем, то состав генотипов потомков будет совпадать с таковым у отобранной группы родителей. Следовательно, и средние значения признаков потомков будут совпадать с их средними значениями у родителей, тем самым величина ответа на отбор сравняется с селекционным дифференциалом. Очевидно, что в данном случае h 2 = 1.
Обычно же ответ на отбор меньше селекционного дифференциала, так как часть особей попадает в отобранную группу совершенно случайно (вследствие счастливого стечения средовых факторов), поэтому особенность их фенотипа наследоваться не может. Ведь для того, чтобы потомки этих «баловней судьбы» (а не генотипа) попали в отбираемую группу, им снова должно повезти. Но случай на то и случай, чтобы не повторяться регулярно.
В генетике количественных признаков доказывается, что наследуемость равна доле генетической дисперсии в полной фенотипической дисперсии признака. Вспоминая, что эта последняя складывается из генетической и средовой компонент, сказанное можно передать формулой
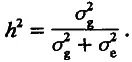 (4.20)
(4.20)
Повторяя процедуру отбора, направленного на изменение величины признака, в течение большого числа поколений мы можем в конце концов добиться фиксации генотипов, включающих в себя только сильные или только слабые аллели (в зависимости от направления отбора). Ясно, что в таком случае весь запас генетической изменчивости будет исчерпан, и дальнейший ответ на отбор станет невозможным.
Теперь обратимся к эксперименту. Действительно, у животных через несколько поколений отбора ответ на него обычно прекращается, и среднее значение признака выходит на плато. Однако интерпретировать это явление не так просто.
Негативные эффекты искусственного отбора
Первый лабораторный эксперимент по отбору на изменение количественных признаков был проведен в начале XX века американским генетиком У. Кастлом. Он ставил перед собой цель — превратить пегих (пятнистых) крыс в одноцветных. В одном варианте отбор был направлен на увеличение площади белых пятен, в другом — в противоположном направлении. Примерно через 20 поколений Кастл был близок к цели, т. е. ему удалось получить почти белых и почти черных крыс, но опыт пришлось прекратить, так как животные захирели и перестали размножаться. Короче говоря, у крыс резко снизились основные показатели приспособленности.
В дальнейшем сходные результаты были получены на большом числе животных объектов — мышах, курах, золотых рыбках, мучных хрущаках и, конечно, на дрозофиле. Везде по мере сдвига самых разных количественных признаков в любом направлении наблюдалось снижение показателей приспособленности: прежде всего жизнеспособности и плодовитости. Популяции как бы сопротивлялись действию искусственного отбора.
Читать дальшеИнтервал:
Закладка:
