Владимир Бердников - Эволюция и прогресс

- Название:Эволюция и прогресс
- Автор:
- Жанр:
- Издательство:Наука
- Год:1991
- Город:Новосибирск
- ISBN:5-02-030062-4
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Владимир Бердников - Эволюция и прогресс краткое содержание
Автор вводит читателя в круг наиболее интригующих вопросов эволюционной биологии. До сих пор эволюционный прогресс остается предметом бурных, даже ожесточенных споров. По существу, всех биологов можно разделить на сторонников и противников идеи этой формы прогресса. Эволюцию живых организмов обычно связывают с ростом их сложности и степени совершенства, однако до сих пор нет строгих критериев этой оценки. Главная мысль, развиваемая автором, состоит в том, что основные атрибуты прогресса — усложнение строения и повышение уровня надклеточной организации — являются лишь следствием постоянно идущего отбора на повышение эволюционной пластичности видов.
Книга предназначена для биологов широкого профиля, а также всех интересующихся вопросами эволюции живых существ.
Эволюция и прогресс - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Заметим, что, введя представление об адаптируемости, мы получили, как следствие, макроэволюционную тенденцию к ее росту. Эту тенденцию можно было бы интерпретировать как накопление эволюционирующей филетической группой информации о неблагоприятных факторах, действующих в ее экологическом пространстве (адаптивной зоне). Как подойти к анализу подобного рода информации, вероятно, запечатленной геномом? Скорее всего такая генетическая запись должна каким-то образом отразиться на фенотипе особей. Поэтому и тенденция к росту адаптируемости в ходе эволюции филетической группы могла бы выглядеть как долговременная тенденция к изменению в определенном направлении морфологии особей ее видов. В связи с этим встает вопрос, не являются ли хорошо известные палеонтологам долговременные тенденции отражением процесса возрастания адаптируемости.
Макроэволюционные тенденции
Крупнейший вклад палеонтологии в наши знания об эволюции живых организмов состоит в обнаружении во многих филетических группах особых очень долговременных тенденций к сдвигу в одном направлении некоторых количественных признаков. Пожалуй, самой известной из них является тенденция к увеличению размеров индивидов — так называемое правило Копа. Значительный эмпирический материал свидетельствует, что обычно зоологические таксоны крупных животных происходят от видов с относительно небольшими особями. Между прочим, род Homo происходит от гораздо более мелких обезьян.
Э. Коп объяснил свое правило как классический ламаркист (кстати, основоположник психоламаркизма). Он считал, что в основе открытой им тенденции лежит пресловутая «ростовая сила», стремящаяся под действием психики животного увеличить его размеры. Такой филогенетический рост тела протекает беспрепятственно в периоды геологического затишья и сопровождается разрастанием многих морфологических структур. Тенденции противостоит процесс вымирания гигантов и монстров при резких сменах климата.
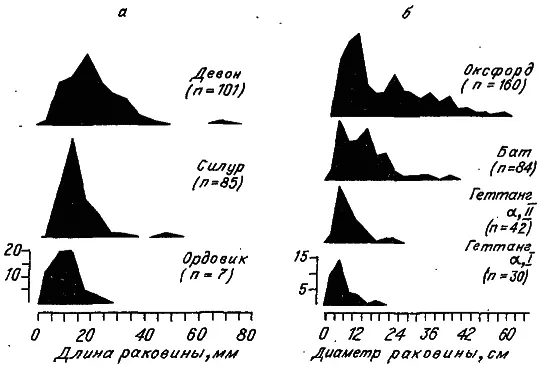
Рис. 17. Изменение видовых распределений брахиопод (а) и аммонитин (б) по размеру раковины в историческом развитии (по: [Stanley, 1973]).
Отметим одну особеннность коповского ламаркизма: внутренние стремления индивида снижают адаптируемость филетической линии и влекут последнюю к гибели. Любопытно, что включение в эволюционный сценарий особых роковых факторов — довольно обычное дело для теоретиков-эволюционистов. Достаточно вспомнить о трех таких драматических историях: 1) самоубийственной тенденции юрских устриц (грифей) к созданию нераскрывающихся раковинных створок; 2) инадаптивном сверхразрастании непомерно тяжелых рогов у ирландского оленя; 3) развитии сверхмощных клыков, затрудняющих кусание, у саблезубых кошек.
Все эти линии на самом деле вымерли, что, конечно, не противоречит предположению об инадаптивном характере данных тенденций. Правда, вымирание филетических линий — дело обычное. Ведь вымерли же, например, все виды семейства Hominidae, за исключением Н. sapiens, и вообще филетический пучок высших приматов не выглядит преуспевающим. Не станем же мы говорить об инадаптивном сверхразрастании головного мозга у гоминид. Кроме того, отметим, что все отмеченные выше случаи «сверхразвития» рабочих структур противоречат принципу симморфоза, согласно которому ни один орган не бывает развит свыше потребности в нем в моменты максимальных нагрузок.
Однако вернемся к правилу Копа. Проследим в какой-нибудь филетической группе за динамикой видовых распределений по величине особей. На рис. 17, а приведены распределения по диаметру раковины у брахиопод отряда Rynchonellida на трех этапах их исторического развития. Аналогичную картину демонстрируют и аммонитины юры (рис. 17, б). Первое, что бросается в глаза, — это увеличение размаха распределений, обусловленное прежде всего появлением видов с большими размерами особей. Кроме того, эти распределения асимметричны — с длинным «хвостом» в правой части. Очевидно, что среднее значение (центр тяжести) распределений также сдвигается с ходом времени вправо. Отметим, что виды с мелкими особями продолжали существовать всегда, хотя их доля постепенно убывала.
Сходную картину можно наблюдать и для видовых распределений по признакам, величина которых хорошо коррелирует с массой тела. Чтобы понять это макроэволюционное явление, сначала рассмотрим, как вообще формируются распределения видов по количественным признакам. Отметим, что после перехода к логарифмической шкале измерения признака многие из этих распределений симметризуются, становясь близкими к нормальным.
Межвидовые дистанции
Мы на глаз, без труда различаем млекопитающих, принадлежащих разным родам. Например, волка и лисицу, рысь и леопарда, козу и овцу и т. д. Еще легче разобраться с представителями разных семейств, отрядов и других таксонов высокого ранга. Сортировка видов одного рода требует большего внимания, хотя в случае млекопитающих она обычно удается, вероятно, помогает собственная принадлежность к данному классу. Видимо, поэтому мы можем отличить друг от друга льва, тигра и леопарда, несмотря на то, что все они входят в один род Panthera. Кстати, отличить череп льва от черепа тигра под силу только специалисту, а гибрид этих самых сильных кошек — тигролев — вполне жизнеспособен. Куда труднее различить родственные виды у насекомых, особенно когда речь идет о так называемых видах-двойниках.
Человек при оценке степени морфологического сходства пользуется присущей ему интуитивной способностью распознавать образы. «Ненаучность» такого метода очевидна. Первая вполне научная система организмов была создана К. Линнеем в 1735 г. Она основана на сопоставлении рядов четко регистрируемых морфологических признаков, которыми можно охарактеризовать типичную особь. Чем больше признаков совпадает в таких рядах, тем выше степень сходства сравниваемых видов. Чем меньше число совпадений, тем выше ранг таксономической дистанции между видами. Слабость этого подхода заключается в субъективном характере выбора признаков, положенных в основу системы.
К сожалению, более объективные молекулярные методы обычно дают информацию только о времени эволюционной дивергенции видов. Дело в том, что удобные для регистрации различия в строении белков или нуклеиновых кислот чаще всего обусловлены нейтральными аллельными заменами, которые фиксируются с более или менее постоянной скоростью — скоростью мутирования (см. (3.26)).
В настоящее время существует ряд способов измерения так называемой морфологической (фенотипической) дистанции между двумя группами особей. Несмотря на различия в математическом оформлении, все эти оценки являются сложным отражением того, во сколько раз (или на сколько процентов) отличаются значения одних и тех же признаков у сравниваемых групп особей. По-видимому, из-за этого оцененные разными способами значения морфологических дистанций неплохо коррелируют друг с другом.
Читать дальшеИнтервал:
Закладка:
