Ник Лейн - Лестница жизни. Десять величайших изобретений эволюции

- Название:Лестница жизни. Десять величайших изобретений эволюции
- Автор:
- Жанр:
- Издательство:Act, corpus
- Год:2014
- Город:Москва
- ISBN:978-5-17-079731-8
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Ник Лейн - Лестница жизни. Десять величайших изобретений эволюции краткое содержание
Как возникла жизнь? Откуда взялась ДНК? Почему мы умираем? В последние десятилетия ученые смогли пролить свет на эти и другие вопросы происхождения и организации жизни. Известный английский биохимик реконструирует историю всего живого, описывая лучшие изобретения эволюции, и рассказывает, как каждое из них, начиная с самой жизни и генов и заканчивая сознанием и смертью, преображало природу нашей планеты и даже саму планету.
Лестница жизни. Десять величайших изобретений эволюции - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
По Дарвину, все живое развивается путем медленного накопления различий, возникающих по мере того, как эволюционные линии расходятся, отделившись от общего предка. Результатом этого процесса оказывается ветвящееся дерево, и нет никаких сомнений, что именно такие деревья лучше всего отображают эволюцию большинства организмов, которых видно невооруженным глазом, то есть, по сути, большинства крупных эукариот. Но так же ясно, что деревья - не лучшие схемы для отображения эволюции микробов, будь то бактерии, архей или одноклеточные эукариоты.
Есть два процесса, вступающие в противоречие с дарвиновскими генеалогическими деревьями: горизонтальный перенос генов и слияние целых геномов. Специалисты по эволюции микробов, пытающиеся разобраться во взаимоотношениях бактерий и архей, с досадной регулярностью сталкиваются с горизонтальным переносом генов. Этот несколько неуклюжий термин означает просто передачу генов от одного организма к другому. В результате геном, который материнская клетка бактерии передает дочерней клетке, может быть и таким же, и не таким, как тот, который сама материнская клетка унаследовала от своей “матери”. Некоторые гены, например исследованный Везе ген рибосомальной РНК, обычно передаются вертикально, из поколения в поколение, в то время как другими клетки микробов, часто совершенно неродственные, постоянно обмениваются3. Общая картина оказывается чем-то средним между деревом и сетью, где ключевые гены (как гены рибосомальной РНК) обычно образуют дерево, а для других характерно образование сети. Вопрос, существует ли такой набор ключевых генов, представителями которого микроорганизмы вообще никогда не обмениваются путем горизонтального переноса, по-прежнему спорен. Если таких генов не бывает, то сама идея проследить за ходом эволюции эукариот вплоть до неких конкретных групп прокариот лишена смысла. Те или иные группы организмов могут иметь определенное происхождение лишь в том случае, если они наследуют признаки непосредственно от собственных предков, а не от каких-то других случайно подвернувшихся групп. Но что если небольшой набор ключевых генов все-таки никогда не передается горизонтально? Что тогда можно сказать о происхождении той или иной группы? Можно ли считать, что кишечная палочка остается кишечной палочкой, если 99 % ее генов случайным образом заменяются другими?4
Со слиянием геномов связаны похожие трудности. Здесь проблема состоит в том, что дарвиновское дерево переворачивается вверх ногами: вместо эволюционного расхождения мы получаем схождение. Тогда возникает вопрос, кто из двух (или большего числа) участников такого схождения отражает истинный ход эволюции? Если проследить наследование одного лишь гена рибосомальной РНК, мы получим обычное ветвящееся дарвиновское древо, но если рассмотреть большое число генов или целые геномы, мы получим что-то вроде кольца, в котором разошедшиеся ветви вновь сходятся и сливаются.
Нет никаких сомнений, что клетки эукариот представляют собой генетические химеры. Данные, свидетельствующие об этом, никто не ставит под сомнение. Вопрос, по поводу которого исследователи теперь разделяются на враждующие фракции, в том, насколько большое значение нужно придавать эволюции по Дарвину, и насколько большое - интенсивным процессам генетического слияния. Иначе говоря, сколько свойств эукариотических клеток возникло путем постепенной эволюции клетки-хозяина, а сколько могло развиться только лишь после генетического слияния. За несколько десятилетий о происхождении эукариотической клетки были выдвинуты десятки теорий, от умозрительных, если не сказать надуманных, до основанных на детальных биохимических реконструкциях. Ни одна из них пока не получила однозначного подтверждения. Все эти теории можно разделить на две большие группы: придающие особое значение постепенному дарвиновскому расхождению и резкому генетическому слиянию. Эти группы соответствуют двум противоборствующим сторонам другого, более давнего спора между биологами: теми, кто доказывает, что эволюция идет путем постепенных непрерывных изменений, и теми, кто настаивает на существовании продолжительных периодов застоя, или равновесия, прерывающихся иногда внезапными впечатляющими изменениями. То есть, как шутили в свое время, споров о том, ползком или прыжками идет эволюция5.
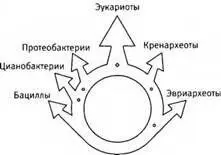
“Кольцо жизни”. Последний общий предок всего живого располагается внизу, где его потомки разделяются на бактерий (слева) и архей (справа). Представители тех и других вновь сливаются вверху, давая начало химерным организмам - эукариотам.
Применительно к эукариотической клетке Кристиан де Дюв назвал представления первой группы гипотезой “примитивного фагоцита”, а второй - гипотезой “судьбоносной встречи”. Идея “примитивного фагоцита” соответствует дарвиновской концепции, и среди ее сторонников выделяются оксфордский эволюционист Том Кавалир-Смит и сам Кристиан де Дюв. В основе этой гипотезы лежит предположение, что предки эукариотических клеток постепенно накопили все признаки, свойственные клеткам современных эукариот: ядро, настоящий половой процесс, клеточный скелет и, что самое важное, способность к фагоцитозу, то есть поглощению других клеток путем изменения формы, окружения их, заглатывания и последующего внутреннего переваривания. Единственной чертой, которой, в отличие от клеток современных эукариот, их предполагаемый непосредственный предок, примитивный фагоцит, не обладал, было наличие митохондрий, которые вырабатывают энергию, используя для этого кислород. Вероятно, ему приходилось получать энергию посредством брожения - гораздо менее эффективного процесса.
Но для фагоцита проглотить предков нынешних митохондрий было тривиальной задачей. Что могло быть проще? Более того, если не путем фагоцитоза, то как могла одна клетка оказаться внутри другой? Обладание митохондриями, разумеется, предоставило примитивному фагоциту важное преимущество (они должны были произвести революцию в его способе получения энергии), но не привело ни к каким принципиальным изменениям его устройства. После того как фагоцит обзавелся митохондриями, он по-прежнему остался фагоцитом, хотя теперь у него появилась возможность получать больше энергии. Однако многие гены из порабощенных митохондрий могли постепенно перейти в ядро и встроиться в геном клетки-хозяина и именно этой передачей объясняется, согласно данной гипотезе, химерная природа современных эукариотических клеток. Их бактериальное наследие составляют гены, взятые у митохондрий. Таким образом, сторонники гипотезы примитивно го фагоцита не оспаривают химерную природу современных эукариот, но предполагают, что предковой для нынешних эукариот клеткой-хозяином, то есть первой настоящей, хотя и примитивной эукариотической клеткой, был нехимерный фагоцит
Читать дальшеИнтервал:
Закладка:



![Уильям Моэм - Десять величайших романов человечества [litres]](/books/1068057/uilyam-moem-desyat-velichajshih-romanov-chelovechestva.webp)
![Ник Лэйн - Вопрос жизни [Энергия, эволюция и происхождение сложности]](/books/1098393/nik-lejn-vopros-zhizni-energiya-evolyuciya-i-proisho.webp)




