Павел Симонов - Эмоциональный мозг

- Название:Эмоциональный мозг
- Автор:
- Жанр:
- Издательство:Питер
- Год:2021
- ISBN:978-5-44613-947-7
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Павел Симонов - Эмоциональный мозг краткое содержание
Эмоциональный мозг - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
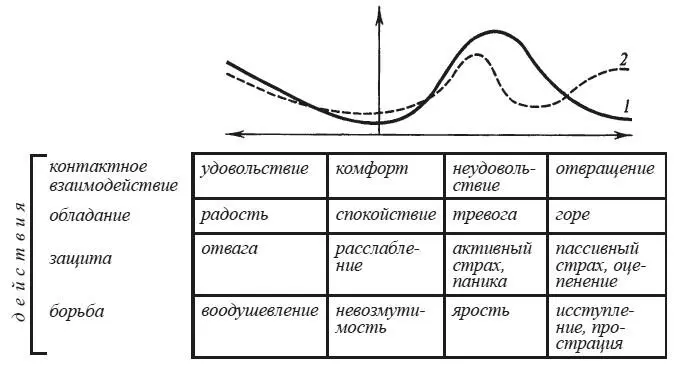
Изменения электроэнцефалограммы при эмоциональных реакциях человека
Степень эмоционального напряжения нередко рассматривают как феномен, идентичный уровню бодрствования. Согласно этим представлениям эмоциональное возбуждение занимает в континууме бодрствования крайнее положение, диаметрально противоположное глубокому медленноволновому сну и коме. Подобный взгляд на природу эмоционального возбуждения, подкрепленный открытием функций ретикулярной формации мозга, получил свое наиболее законченное воплощение в «активационной теории эмоций» Дональда Б. Линдсли [1960]. Однако факты демонстрируют многочисленные примеры явной диссоциации между степенью эмоционального возбуждения, если о нем судить по сдвигам вегетативных функций, и уровнем бодрствования, диагностированным по суммарной электрической активности мозга. Ожидание болевых раздражений в опытах Р. Фроста с соавторами приводило к сдвигам частоты сердцебиений и электросопротивления кожи без достоверных изменений альфа-ритма электроэнцефалограммы — ЭЭГ [Frost, Burish, Holmes, 1978]. В состоянии страха, при восприятии эмоционально окрашенных слов, во время сдачи экзаменов были зарегистрированы не угнетение, а усиление альфа-ритма, повышение его амплитуды, возрастание альфа-индекса.
Вопрос о соотношении вегетативных и ЭЭГ признаков эмоционального возбуждения был подвергнут систематическому анализу в экспериментах М. Н. Русаловой [1979]. На рис. 15 хорошо видно, что эмоциональное напряжение, о котором судили по учащению сердцебиений, может сопровождаться у одного и того же субъекта как угнетением, так и экзальтацией альфа-ритма. Чем различаются эти две ситуации? В первом случае субъект ожидал реальное болевое раздражение кожи предплечья. Во втором — ему была дана инструкция мысленно представить болевое раздражение в конце падающего отсчета, записанного на магнитофон.
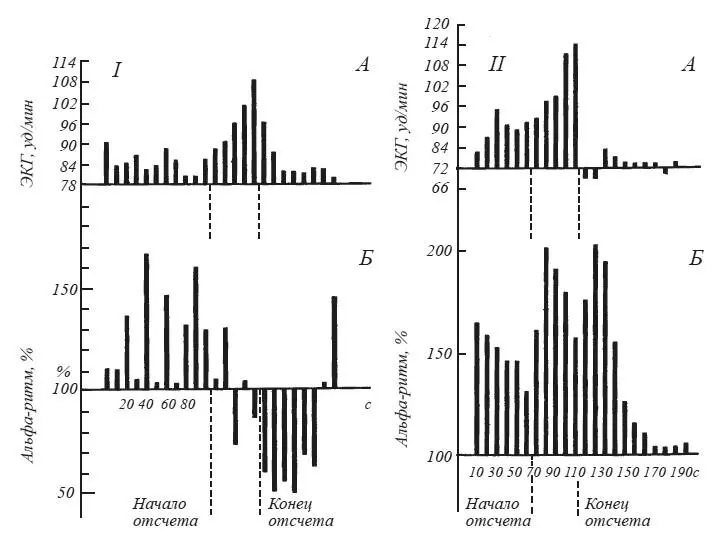
Рис. 15.Динамика изменений амплитуды (Б) альфа-ритма (по показанию пера интегратора) и частоты сердечного ритма (Д) после инструкции, в период отсчета и после отсчета
Альфа-ритм (11 Гц) выражен в процентах к фону, принятому за 100; I — действительное ожидание боли; II — воображаемое ожидание боли (по М. Н. Русаловой).
Выше мы говорили, что эмоциональная реакция может возникнуть как при поступлении информации из внешней среды, так и при извлечении следов ранее полученных впечатлений из памяти. Многочисленные эксперименты, поставленные М. Н. Русаловой (Валуевой), равно как и данные, почерпнутые в литературе, убеждают в том, что депрессия альфа-ритма характерна для ситуаций, в которых внимание субъекта обращено к окружающей его внешней среде. Если субъект преимущественно сосредоточен на следах, хранящихся в памяти, даже высокая степень эмоционального возбуждения не ведет к угнетению альфа-ритма, а зачастую сопровождается его экзальтацией. Подобный вывод хорошо согласуется с представлением об альфа-ритме как электрофизиологическом корреляте механизма, квантующего поток поступающей в мозг афферентации [Walter, 1954]. Нуну и Осселтон показали, что короткие зрительные стимулы не воспринимаются, если они подаются во время определенных фаз альфа-ритма. По мнению авторов, альфа-ритм коррелирует с деятельностью «нервной задвижки», которая периодически прерывает поступление информации в мозг [Nunn, Osselton, 1974].
Может возникнуть вопрос: почему же при сильном потоке афферентации, когда необходимость в работе «задвижки» особенно велика, альфа-ритм исчезает, депрессируется? Здесь не следует отождествлять деятельность механизма квантования с выраженностью его электрофизиологических коррелятов. Хорошо регистрируемый альфа-ритм при закрытых глазах, тишине и покое — это «холостой ход» квантующего механизма, который, кстати, существует и у лиц без выраженного альфа-ритма в ЭЭГ. Сказанное справедливо и для динамики тета-ритма, о котором сейчас пойдет речь.
Одним из ЭЭГ, симптомов эмоционального возбуждения служит усиление тета-ритма с частотой колебаний 4–7 Гц. С помощью анализаторов частотного спектра ЭЭГ тета-ритм был зарегистрирован у здоровых людей при отрицательных [Суворова, 1975] и положительных [Walter, 1953; Валуева, 1967] эмоциях. У детей, занятых деятельностью, окрашенной положительными эмоциями, тета-ритм лучше выражен в правом полушарии большого мозга, особенно в лобных его областях [Денисова, 1978]. Для понимания функционального значения тета-ритма интересны данные Брауна [Brown, 1971], который демонстрировал субъекту альфа-, бета- и тета-полосы его собственной ЭЭГ в виде различной цветовой окраски экрана. Испытуемых спрашивали, какому их субъективному состоянию соответствует тот или иной цвет. Оказалось, что для тета-ритма характерны обдумывание планов, неопределенность мысленно анализируемой ситуации, изменение темы размышлений, решение технических и житейских проблем, «сны наяву». Нарастание процентного содержания тета-ритма в спектре ЭЭГ совпадает со снижением бдительности человека-оператора к внешним сигналам [Beatty, Greenberg, Deibler, O’Hanlon, 1974].
Для того чтобы оценить функциональное значение изменений ЭЭГ при эмоциях, остановимся на гипотетическом механизме ритмических колебаний биопотенциалов мозга. Согласно гипотезе П. Андерсена и Дж. Экклса, важную роль в формировании этих колебаний играют тормозные нейроны [Andersen, Eccles, 1962]. Это дает основание применить к взаимодействию возбужденных и тормозящих их деятельность нервных микроструктур теорему В. Вольтерра [1976] о закономерных изменениях амплитуды и частоты осцилляций в системе, состоящей из двух типов функционально антагонистических элементов (табл. 3).
Таблица 3.Изменение электроэнцефалограммы, предсказываемое теоремой Вольтерра
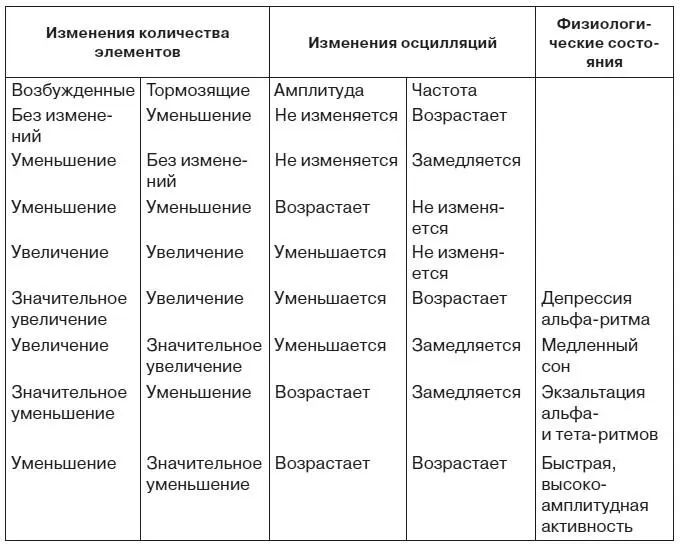
В какой мере картина, представленная в таблице, совпадает с физиологическими фактами? Наиболее бесспорен, пожалуй, механизм классической депрессии альфа-ритма, которая определяется нарастанием числа возбужденных и тормозящих элементов с преимущественным увеличением первых из них. Весьма неожиданным может показаться тот факт, что картина, характерная для медленного сна, также обусловлена увеличением возбужденных и тормозящих элементов, только число последних нарастает в большей мере, чем количество первых. Впрочем, данные последних лет не позволяют отвергнуть этот вывод. Усиление активности нейронов коры и ретикулярной формации во время сна побуждает рассматривать сон скорее как результат перестройки взаимодействия нервных элементов, чем как состояние «разлитого торможения» коры и подкорки в старом понимании этого термина. Ряд авторов придают важное значение нарастанию активности механизмов возвратного торможения в генезе медленного сна.
Читать дальшеИнтервал:
Закладка: