Кирилл Еськов - История земли и жизни на ней

- Название:История земли и жизни на ней
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Кирилл Еськов - История земли и жизни на ней краткое содержание
Экспериментальное учебное пособие для старших классов.
"Авторское предуведомление
Цель настоящего учебного курса мне видится в том, чтобы у ученика возникла максимально целостная картина функционирования биосферы Земли в процессе ее исторического развития. Между тем одно из положений теории систем гласит: систему невозможно оптимизировать по двум независимым параметрам одновременно; в частности, добиваясь целостности рисуемой картины, неизбежно приходится жертвовать ее детальностью, или наоборот. Попытка запихнуть в голову ученика побольше конкретных фактов в отсутствие некоей обобщающей концепции неизбежно приведет нас к созданию ухудшенной копии старого университетского курса палеонтологии – унылого мартиролога вымерших организмов, который по сдаче экзамена следует забыть, как страшный сон. Именно поэтому во многих случаях я вполне сознательно жертвовал палеонтологической и геологической конкретикой в пользу теоретических (иногда к тому же – в достаточной степени умозрительных) обобщений".
История земли и жизни на ней - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
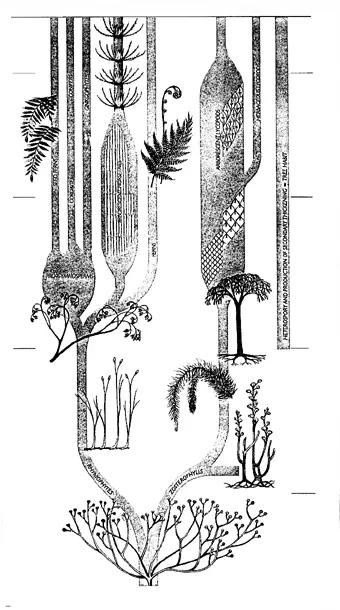
РИСУНОК 30. Родословное древо первых наземных растений.
Появление сосудистых растений с их жесткими вертикальными осями вызвало целый каскад экосистемных новаций, изменивших облик всей биосферы:
1. Фотосинтезирующие структуры стали располагаться в трехмерном пространстве, а не на плоскости (как это было до сих пор – в период господства водорослевых корок и лишайников). Это резко увеличило интенсивность образования органического вещества и, тем самым, суммарную продуктивность биосферы.
2. Вертикальное расположение стволов сделало растения более устойчивыми к занесению смываемым мелкоземом (по сравнению, например, с водорослевыми корками). Это уменьшило безвозвратные потери экосистемой неокисленного углерода (в виде органики) – совершенствование углеродного цикла.
3. Вертикальные стволы наземных растений должны быть достаточно жесткими (по сравнению с водными макрофитами). Для обеспечения этой жесткости возникла новая ткань – древесина, которая после гибели растения разлагается относительно медленно. Таким образом, углеродный цикл экосистемы обретает дополнительное резервное депо и, соответственно, стабилизируется.
4. Появление постоянно существующего запаса трудноразложимой органики (сконцентрированного в основном в почве) ведет к радикальной перестройке пищевых цепей. С этого времени большая часть вещества и энергии оборачивается через детритные, а не через пастбищные цепи (как это было в водных экосистемах).
5. Для разложения трудноусваиваемых веществ, из которых состоит древесина – целлюлозы и лигнина – потребовались новые типы разрушителей мертвой органики. С этого времени на суше роль основных деструкторов переходит от бактерий к грибам.
6. Для поддержания ствола в вертикальном положении (в условиях действия силы тяжести и ветров) возникла развитая корневая система: ризоидами – как у водорослей и мохообразных – тут уже не обойдешься. Это привело к заметному снижению эрозии и появлению закрепленных (ризосферных) почв.
С.В. Мейен полагает, что суша должна была покрыться растительностью к концу девона (зигенский век) [36] К концу девона уже появились представители почти всех основных отделов высших растений, не только споровых (плауновидные, папоротникообразные, хвощеобразные), но и голосеменных (гинкговые).
, поскольку с начала следующего, карбонового, периода на Земле образуются уже практически все типы осадков, отлагающихся ныне на континентах. В дозигенские же времена континентальные осадки практически отсутствуют – видимо, в связи с их постоянным вторичным размывом в результате нерегулируемого стока. В самом начале карбона на континентах начинается угленакопление – а это свидетельствует о том, что на пути стока вод стояли мощные растительные фильтры. Не будь их, остатки растений непрерывно смешивались бы с песком и глиной, так что получались бы обломочные породы, обогащенные растительными остатками – углистые сланцы и углистые песчаники, а не настоящие угли.
Итак, возникшая в прибрежных амфибиотических ландшафтах густая «щетка» из гелофитов (можно назвать ее «риниофитный камыш») начинает действовать как фильтр, регулирующий плащевой сток: она интенсивно отцеживает (и осаживает) сносимый с суши обломочный материал и формирует тем самым стабильную береговую линию. Некоторым аналогом этого процесса может служить формирование крокодилами «аллигаторовых прудов»: животные постоянно углубляют и расширяют населяемые ими болотные водоемы, выбрасывая грунт на берег. В результате их многолетней «ирригационной деятельности» болото превращается в систему чистых глубоких прудов, разделенных широкими облесенными «дамбами». Так и сосудистая растительность в девоне разделила пресловутые амфибиотические ландшафты на «настоящую сушу» и «настоящие пресноводные водоемы». Не будет ошибкой сказать, что именно сосудистая растительность стала истинным исполнителем заклинания: «Да будет твердь!» – отделивши оную твердь от хляби...
Именно с вновь возникшими пресноводными водоемами связано и появление в позднем девоне (фаменский век) первых тетрапод (четвероногих) – группы позвоночных, имеющих две пары конечностей; она объединяет в своем составе амфибий, рептилий, млекопитающих и птиц (попросту говоря, тетраподы – это все позвоночные, кроме рыб и рыбообразных). В настоящее время общепринято, что тетраподы ведут свое происхождение от кистеперых рыб (Rhipidistia) (рисунок 31); эта реликтовая группа имеет ныне единственного живого представителя, латимерию [37] Широко известна истинно романтическая история открытия этого «живого ископаемого», описанная в замечательной книге Дж.Смита «Старина четвероног». Следует, правда, заметить, что образ жизни латимерии не имеет ничего общего с тем, что вели девонские рипидистии: она живет в Индийском океане на глубинах в несколько сот метров.
. Достаточно популярная некогда гипотеза происхождения четвероногих от другой реликтовой группы рыб – двоякодышащих (Dipnoi), ныне практически не имеет сторонников.
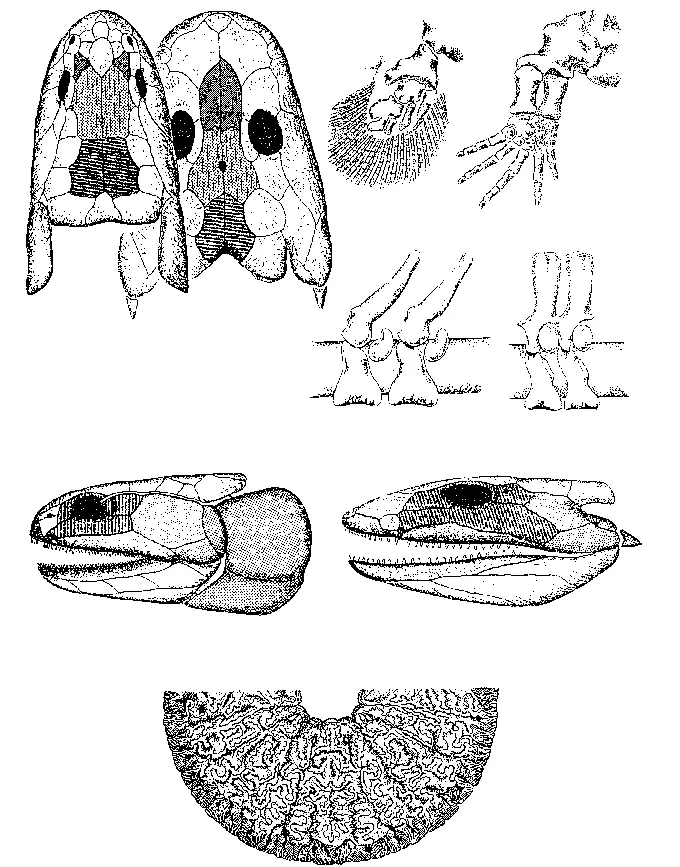
РИСУНОК 31. Преемственность анатомических структур (череп, конечности и позвонки) кистеперых рыб и амфибий: идентичные кости черепа имеют одинаковую заливку; идентичные кости обозначены одинаковыми буквами; отдельно – поперечный шлиф половинки зуба лабиритнтодонта.
Необходимо отметить, что в прежние годы возникновение ключевого признака тетрапод – двух пар пятипалых конечностей – считали их однозначной адаптацией к наземному (или по меньшей мере амфибиотическому) образу жизни. Ныне, однако, большинство исследователей склоняется к тому, что «проблема появления четвероногих» и «проблема их выхода на сушу» – суть вещи разные и даже не связанные между собою прямой причинной связью. Предки тетрапод жили в мелководных, часто пересыхающих, обильно заросших растительностью водоемах непостоянной конфигурации. Судя по всему, конечности появились для того, чтобы передвигаться по дну водоемов (это особенно важно, когда водоем обмелел настолько, что у тебя уже спина начинает вытарчивать наружу) и продираться сквозь плотные заросли гелофитов; особенно же полезны конечности оказались для того, чтобы при пересыхании водоема переползти посуху в другой, соседний.
Первые, девонские, тетраподы – примитивные амфибии лабиринтодонты [38] Старое название, которое вы можете встретить в книгах – стегоцефалы – сейчас не употребляют.
(название это происходит от их зубов с лабиринтоподобными складками эмали – структура, прямо унаследованная от кистеперых: см. рисунок 31), такие как ихтиостега и акантостега, в захоронениях всегда встречаются вместе с рыбами, которыми, судя по всему, и питались. Они были покрыты чешуей, как рыбы, имели хвостовой плавник (похожий на тот, что мы видим у сома или налима), органы боковой линии и – в некоторых случаях – развитый жаберный аппарат; конечность их еще не пятипалая (число пальцев достигает 8), и по типу сочленения с осевым скелетом – типично плавательная, а не опорная. Все это не оставляет сомнения в том, что существа эти были чисто водными (рисунок 32); если они и появлялись на суше при неких «пожарных» обстоятельствах (пересыхание водоема), то компонентом наземных экосистем наверняка не являлись [39] Не называем же мы «наземным существом» угря, который способен ночью, по росистой траве, переползти из одного водоема в другой, преодолевая расстояние в несколько сот метров!
. Лишь много позже, в карбоновом периоде, появились мелкие наземные амфибии – антракозавры, которые, судя по всему, питались членистоногими, но об этом речь впереди (см. главу 10).
Интервал:
Закладка:




