Кирилл Еськов - История земли и жизни на ней

- Название:История земли и жизни на ней
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Кирилл Еськов - История земли и жизни на ней краткое содержание
Экспериментальное учебное пособие для старших классов.
"Авторское предуведомление
Цель настоящего учебного курса мне видится в том, чтобы у ученика возникла максимально целостная картина функционирования биосферы Земли в процессе ее исторического развития. Между тем одно из положений теории систем гласит: систему невозможно оптимизировать по двум независимым параметрам одновременно; в частности, добиваясь целостности рисуемой картины, неизбежно приходится жертвовать ее детальностью, или наоборот. Попытка запихнуть в голову ученика побольше конкретных фактов в отсутствие некоей обобщающей концепции неизбежно приведет нас к созданию ухудшенной копии старого университетского курса палеонтологии – унылого мартиролога вымерших организмов, который по сдаче экзамена следует забыть, как страшный сон. Именно поэтому во многих случаях я вполне сознательно жертвовал палеонтологической и геологической конкретикой в пользу теоретических (иногда к тому же – в достаточной степени умозрительных) обобщений".
История земли и жизни на ней - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
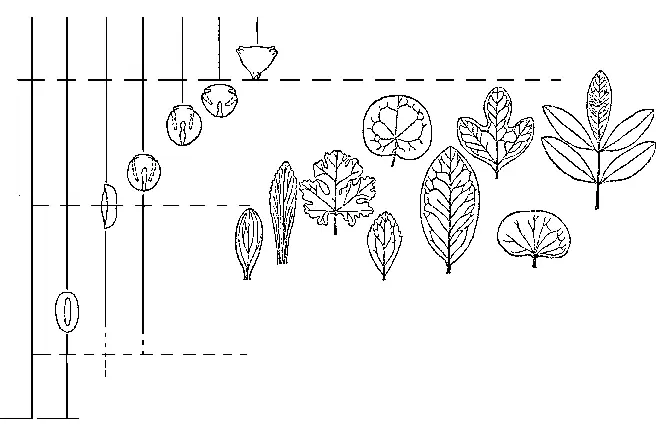
РИСУНОК 50. Распределение в меловых отложениях основных типов пыльцы и листьев покрытосеменных.
В 70-х годах советские палеоэнтомологи исследовали множество местонахождений меловых насекомых в Сибири и Монголии, «закрасив» тем самым самое крупное «белое пятно» в геологической истории класса. Оказалось, что у насекомых график, который отражает долю вымерших таксонов в фаунах разных времен, ведет себя довольно неожиданным образом (рисунок 51). Доля вымерших семейств мало меняется на протяжении большей части мезозоя, и на границе юры и мела составляет около 60% ; затем, в течении раннего мела, доля эта стремительно падает от 60 до 20% , в позднем мелу это падение резко замедляется (от 20 до 5%), а к началу кайнозоя кривая вновь «выходит на плато», только уже в районе 5%. Иными словами, у насекомых самая резкая фаунистическая смена, связанная с массовым вымиранием мезозойских групп, приходится не на конец мела, а на его середину – в точности как у растений. На этом основании В.В. Жерихин (1978) предложил следующую модель среднемелового биоценотического кризиса.
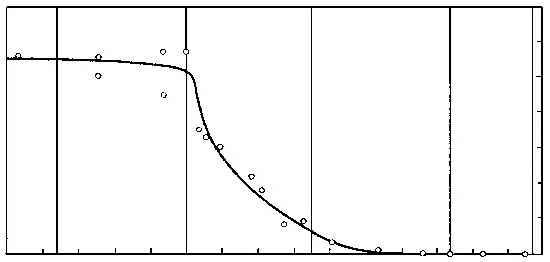
РИСУНОК 51. Эволюция насекомых в мезозое: процент вымерших семейств от общего числа семейств насекомых в позднемезозойских и кайнозойских фаунах.
Покрытосеменные возникли в начале мелового периода, первоначально как ценофобы – растения, не входящие в закономерные, сложившиеся в длительной коэволюции сукцессионные ряды [69] Под сукцессией здесь подразумевается не всякая смена растительности, а то, что С.М.Разумовский (1982) называет экогенетической сукцессией: жестко детерменированная последовательность растительных ассоциаций (сериальный ряд), каждая из которых видоизменяет состав почвы так, что он становится более благоприятен для ассоциации-последователя, нежели для нее самой, и приводит в конечном итоге к ассоциации-климаксу, в почвах которой достигнут «нулевой баланс» по органике; климаксная ассоциация не имеет внутренних причин к смене и может существовать неопределенно долго – пока не будет разрушена извне (например, лесным пожаром). Пример экогенетической сукцессии для Подмосковья – зарастание гари иван-чаем (пионерная стадия), березняк, смена его ельником и, далее, дубравой (климаксом); цикл этот длится около 300 лет. Совокупность климакса с ведущими к нему сериальными рядами называют сукцессионной системой; ее можно рассматривать как гомеостат, который имеет равновесное состояние (климакс), а будучи же выведен из него внешними силами, возвращается в исходное положение при помощи ряда стандартизованных реакций (экогенетических смен).
. Они росли в качестве «сорной» растительности по свободным от других растений участкам (на речных отмелях, береговых оползнях, гарях), которые соседствовали с гораздо более обширными участками, занятыми зрелыми сообществами мезофитной растительности. Покрытосеменные были исходно энтомофильными и смогли сформироваться лишь в окружении энтомофильных же голосеменных (таких, как беннетиты, саговники, кейтониевые), используя уже сложившийся к тому времени комплекс их опылителей. Но если покрытосеменные оказались среди пионерных растений мезофита единственными энтомофилами (а, судя по всему, так оно и было), то это должно было дать им гиганские преимущества: именно для локально распределенных, не образующих сплошных зарослей пионеров насекомоопыление особенно выгодно; впрочем, возможно, имело место и прямое конкурентное вытеснение покрытосеменными энтомофильных голосеменных из пионерных сообществ – за счет более эффективного привлечения насекомых, лучшей защиты от повреждения завязи опылителями и т.д.
Основой стремительной среднемеловой экспансии цветковых стало то, что им удалось закрепиться в качестве нормальной пионерной растительности – для чего решающими факторами стали их исходная энтомофильность и наличие среди них травянистых форм (что во много раз убыстряет зарастание поврежденных участков). Покрытосеменные не пытались потеснить всю мезофитную растительность, что было абсолютно нереально; они «всего-навсего» конкурентно вытеснили прежних пионеров – и тем самым блокировали все последующие стадии мезофитной экогенетической сукцессии. Разрушение существовавших в то время закономерных сукцессионных рядов (попросту говоря – мезофитная растительность продолжала существовать там, где она существует, но потеряла способность восстанавливаться после экзогенных нарушений), вызвало полный развал мезозойских наземных экосистем и массовое вымирание входивших в них животных; наиболее интенсивно эта деструкция шла в альбе (захватывая конец апта и начало сеномана).
В.В. Жерихин (1978) обратил внимание на любопытнейшую черту энтомофаун этого времени: многие семейства, характерные для мезозоя, обнаруживаются в них в последний раз, тогда как сколько-нибудь заметного появления новых групп не отмечается, и при этом появляется целый ряд представителей реликтовых для того времени (раннемезозойских и даже позднепалеозойских) групп. Это «всплывание реликтов» считают одним из наиболее ярких симптомов кризиса, охватившего среднемеловые сообщества: вымирание доминирующих раннемеловых групп (наиболее тесно связанных с существующей сукцессионной системой) создает экологический вакуум, который ненадолго заполняют древние формы, оттесненные к тому времени «на обочину жизни».
Складывающиеся на совершенно новой основе раннекайнофитные сообщества первоначально были несбалансированными и неустойчивыми. Слабость биоценотической регуляции и обилие незанятых экологических ниш вели к очень быстрой, взрывной, эволюции различных групп организмов (из насекомых здесь можно отметить высших бабочек и мух, паразитических перепончатокрылых, а также общественных насекомых – муравьев и термитов), по мере же насыщения биоценозов темпы эволюции снижались. Сложные трофические цепи в новых кайнофитных сообществах сформировались очень быстро (листья, поврежденные минирующими бабочками, сами эти бабочки и их высокоспециализированные паразиты из числа наездников Braconidae появляются в летописи почти одновременно, в сеномане) и в дальнейшем менялись мало. «По экологическому облику и систематическому составу , – заключает Жерихин (1980), – позднемеловые фауны значительно сильнее отличаются от раннемеловых, чем от кайнозойских, и могут рассматриваться как ранний, первоначально сильно обедненный, вариант этих последних. В сущности, по набору жизненных форм они имеют не так уж много отличий от современных».
Читать дальшеИнтервал:
Закладка:




