Марк Мосевицкий - Распространенность жизни и уникальность разума?

- Название:Распространенность жизни и уникальность разума?
- Автор:
- Жанр:
- Издательство:ПИЯФ РАН
- Год:2008
- Город:Санкт-Петербург
- ISBN:5-86763-203-2
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Марк Мосевицкий - Распространенность жизни и уникальность разума? краткое содержание
В книге рассматривается возникновение и развитие жизни на Земле. Исследуется влияние на жизнь различных эволюционных катастроф: падение астероидов, глобальные оледенения и т. п. Подчеркивается отсутствие признаков существования других цивилизаций в доступном для общения космосе и как следствие низкая вероятность выхода эволюции на путь, ведущий к разуму.
Распространенность жизни и уникальность разума? - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Выше, говоря о горизонтальном переносе, мы имели в виду передачу отдельных признаков (генов). Такие переносы, постепенно накапливаясь, могли коренным образом преобразовать генетический аппарат и физиологию клетки. Было высказано предположение, что особенно эффективно они происходили при захвате и использовании одних клеток другими в качестве пищи. В частности, именно “пищевыми“ переносами Дулиттл объяснял присутствие большого числа бактериальных генов в хромосомах эукариотов (Doolittle, 1998).
Однако значение, которое невозможно переоценить, сыграла другая форма горизонтального переноса, когда объединялись в единый организм целые клетки.
5.3.1 Концепция формирования митохондрий и хлоропластов путем симбиоза бактериальной клетки и раннего эукариота
Около 2 млрд лет тому назад на Земле создалась критическая для дальнейшего развития жизни ситуация. Фотосинтезирующие бактерии, размножившись, стали производить все больше кислорода. Первоначально, пока на поверхности Земли было много чистых металлов, а в атмосфере присутствовали метан, сероводород и другие восстановленные соединения, практически весь кислород расходовался на их окисление. Однако к указанному сроку практически все выходившие на поверхность металлы были превращены в окислы и в атмосфере начал накапливаться свободный кислород. Для хемотрофных анаэробов, какими были тогда все архе, кислород оказался ядом. Поэтому они были либо обречены на гибель, либо должны были ограничиться нишами, куда кислород не проникал (глубинные области морей и океанов, выходы лавы и подземных вод и др.). Однако у бактерий успели появиться виды (протеобактерии), которые, обзаведясь механизмом окислительного фосфорилирования, стали использовать кислород для получения энергии, запасаемой в АТФ (как при фотосинтезе). Естественно, для них кислород стал не ядом, а благом. Анаэробы могли облегчить свое существование, вступив в состояние симбиоза с протеобактериями, которые, поглощая кислород, осуществляли локальную детоксикацию. Если вернуться к представлению о эукариотах как о продолжении одной из линий архе, то ранние эукариоты были анаэробами и так же, как архе, нуждались в тесном общении с утилизирующими кислород бактериями.
Около 20-ти лет тому назад исследование митохондрий и хлоропластов, присутствующих в клетках животных (только митохондрии) и растений (митохондрии и хлоропласты), неопровержимо доказало их бактериальное происхождение. Тогда же получила признание гипотеза формирования современных эукариот, благодаря имевшим место случаям симбиоза ранних эукариот с бактериями, которые трансформировались в митохондрии (Gray, 1989; Cavalier-Smith, 2002). Митохондрии стали внутриклеточными энергетическими фабриками эукариотической клетки, в которых осуществляется синтез АТФ по механизму аэробного окислительного фосфорилирования (Рис. 5А). На основании генетического анализа разных форм современных эукариот многие исследователи пришли к заключению о единственном эпизоде симбиоза раннего эукариота с П-протеобактерией, в результате которого эукариоты приобрели митохондрии. Симбиоз уже содержавших митохондрии эукариотических клеток с фотосинтезирующими бактериями, которые трансформировались в хлоропласты, дал миру растения, в первую очередь водоросли. Произошло это около 1.5 млрд лет тому назад (Yoon et al., 2004). У различных водорослей, помимо других отличий, обнаружены разные формы хлорофилла, характерные для определенных фотосинтезирующих бактерий. Это означает, что акты симбиоза, приводившие к образованию хлоропластов, были неоднократны. В частности, зеленые водоросли, давшие начало наземным растениям, произошли от эукариотической клетки, вступившей в симбиоз с зеленой бактерией (для обзора Малахов, 2004).
У современных эукариотических организмов некоторые гены, контролирующие работу митохондрий, сохранились в самих органеллах, в которые трансформировались бактерии, а другие переместились в ядро и, функционируя в составе генома эукариотической клетки, продолжают обслуживать своих бывших хозяев. Потому, несмотря на ядерную локализацию, их именуют митохондриальными. К ним относятся гены митохондриальных транспортных РНК, рибосомных РНК и рибосомных белков. Продукты этих генов транспортируются в митохондрии, где участвуют в формировании рибосом бактериального типа для синтеза собственных белков. Поэтому рибосомы митохондрий даже визуально отличаются от рибосом самой эукариотической клетки, унаследованных от архе. В принципе, также обстоят дела и с хлоропластами растений.
5.3.2. Гипотезы, связывающие появление эукариот с актом симбиоза архе и протеобактерии
В последние годы появились основания к пересмотру некоторых элементов эволюционной истории эукариот. Главным образом, изменения касаются самого происхождения эукариотической клетки и ранних этапов ее эволюции.
Преобладающим стало мнение, что клеточной линии, отделившейся от прокариот и самостоятельно трансформировавшейся в ранних эукариот, вообще не существовало. Анализ расшифрованных геномов эукариот и прокариотических организмов, как архе, так и бактерий, показал, что первичные эукариоты возникли не в результате обусловленного мутациями вертикального процесса, а как следствие горизонтального процесса — симбиоза бактерии и архе (Gupta, 1998; Martin and Muller, 1998; Ribeiro and Golding, 1998; Vellai and Vida, 1999; Margulis et al., 2000; Horiike et al., 2002, 2004; Aguilar et al., 2004; Rivera and Lake, 2004; Simonson et al., 2005; Zhaxybayeva et al., 2006). Рассматриваются существенно различные схемы такого симбиоза.
Согласно предложенной рядом авторов гипотезе (Gupta, 1998; Moreira and Lopez-Garcia, 1998; Horiike et al., 2002, 2004), эукариотическая клетка образовалась единственный раз за всю историю Земли при событии симбиоза прокариотической клетки, принадлежащей ветви архе (эоцит) и грамотрицательной бактерии (протеобактерия) (Рис. 5Б). Эти авторы полагают, что клетка архе была поглощена бактерией, образовав ядро композита, а сама бактерия сформировала окружение ядра (цитоплазму). Не исключено, что в симбиоз вступили протопласты или сферопласты, т. е. клеточные формы, лишенные оболочки (Sinkovics J.G., 2001; Wachtershauser G., 2003). Позднее хромосома бактерии переместилась в ядро, сформировав там вместе с хромосомой архе комплексный (химерный) геном эукариотической клетки. В этом геноме “информационные” гены, контролирующие
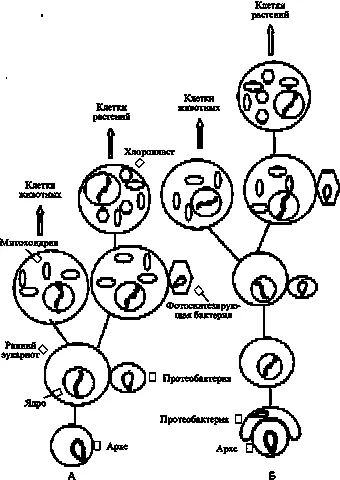
Рис. 5. Формирование современных эукариотических клеток. А. На базе возникших эволюционным путем ранних эукариотов. Б. На базе эукариота, возникшего благодаря акту симбиоза протеобактерии и клетки архе (подробности в тексте). Рисунок создан при участии О. М. Горбенко.
Читать дальшеИнтервал:
Закладка:







