Феликс Филатов - Клеймо создателя

- Название:Клеймо создателя
- Автор:
- Жанр:
- Издательство:Литагент «Ридеро»78ecf724-fc53-11e3-871d-0025905a0812
- Год:неизвестен
- ISBN:978-5-4474-2574-6
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Феликс Филатов - Клеймо создателя краткое содержание
Книга доктора биологических наук Ф. П. Филатова «Клеймо создателя» посвящена одной из версий происхождения жизни на Земле, аргументированной формальными особенностями генетического кода. Эта версия может оказаться также решением парадокса Ферми о существовании разумной жизни за пределами нашей звездной системы. Книга располагает к фундаментальным размышлениям, и ее с интересом прочтут биологи, математики, философы – все, кто интересуется космологией вообще.
Клеймо создателя - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
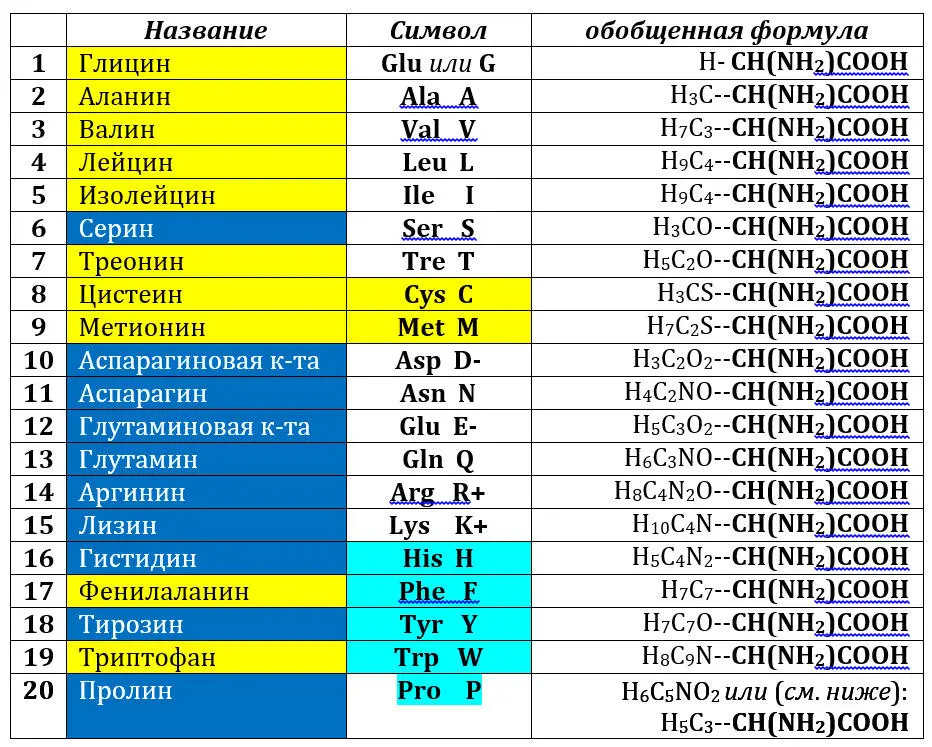
Для обозначения аминокислот (напомним, что кодируемыми являются альфа-L-аминокислоты) используют либо трех-, либо однобуквенные символы; мы – как уже сказано – будем пользоваться последними. В таблице выделены гидрофильные (синие ячейки и белые буквы названий) и гидрофобные (желтые ячейки) аминокислоты, аминокислоты, способные нести заряд, отмечены знаками (+) или (-), ароматические аминокислоты (бирюзовые ячейки в колонке символов, иминокислота пролин – бирюзовое выделение); серусодержащие аминокислоты (желтые ячейки в колонке символов). В формуле молекул справа – одна и та же константная часть (участвующая в пептидной связи; полужирный шрифт), слева – боковая часть молекулы или радикал (R). Молекула пролина приведена к общей схеме гипотетическим размыканием (релаксацией) иминного кольца.
Очевидно, что аминокислоты отличаются друг от друга химической природой боковой цепи, которая состоит из группы атомов в молекуле аминокислоты, связанной с α-углеродным атомом и не участвующей в образовании пептидной связи при синтезе белка. Всѐ разнообразие особенностей структуры и функции белковых молекул связано с химической природой и физико-химическими свойствами радикалов аминокислот. Именно благодаря им, белки наделены рядом уникальных функций, не свойственных другим биополимерам, и обладают химической индивидуальностью. Благодаря им, вновь синтезирующаяся полипептидная цепочка приобретает вторичную структуру, образуя определенной длины однотипные спирали, складчатость и повороты (изломы). Эта структура, в свою очередь, складывается в уникальную третичную, которая и обладает определенными функциями. Они могут быть усилены или модифицированы четвертичной белковой структурой, которую формируют уже не отдельные полипептиды, а их комбинация.
Это общие сведения ( trivia ) о компонентах генетического кода. Приводим его стандартную (каноническую) таблицу. Синим выделены в ней кодирующие и кодируемые элементы группы вырожденности IVоктета 1 (см. ниже); серым – элементы октета 2 групп вырожденности I (темно-серые), III(светлее) и II(еще светлее). В неокрашенных ячейках – терминирующие кодоны. Чтобы подчеркнуть характер непосредственных участников процесса декодирования, то есть молекул РНК, четырьмя основаниями в таблице часто выбираются основания U , C , A и G . Именно на таком порядке настаивал Френсис Крик – не помню точно, почему, – может быть, потому, что, скажем, теория граничных орбиталей химической реактивности, которая была разработана для сравнения вероятностей стабилизации избыточных электронов для различных ДНК-составляющих, предсказывает снижение электронного сродства и потенциалов ионизации, подтверждаемое экспериментальными данными, именно в порядке TCАG 51.
ВТОРОЙ НУКЛЕОТИД
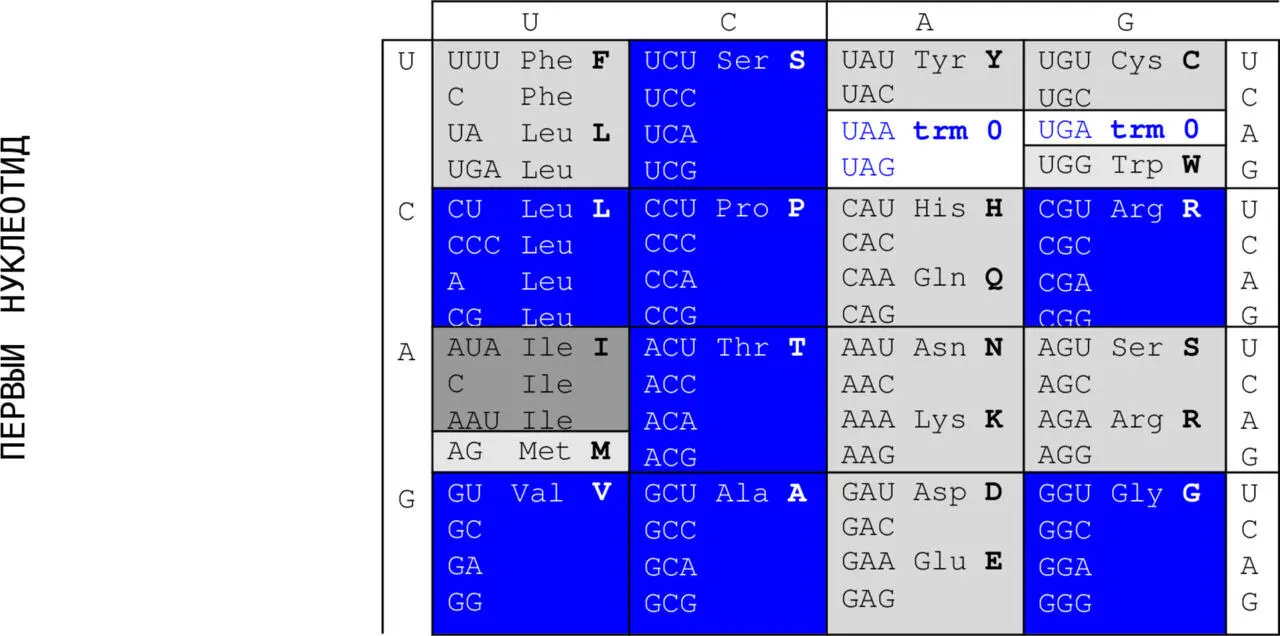
Функция молекулы ДНК – не декодирование, но хранение генетической информации. Поскольку выбор «главной» из этих двух функций – задача очевидно нелепая, таблицы кода с ДНК-основаниями Т , C , A и G встречаются не менее часто.
Напомним теперь восемь основных свойств генетического кода, определяющих сопоставление нуклеотидов и аминокислот, и девятое – альтернативное.
Первое из них – триплетность, означающая, что каждую используемую в живых структурах аминокислоту кодируют три последовательно (то есть от 5`– к 3`-концу цепи) расположенных азотистых основания. Их называют триплетом или кодоном. В таблице первым двум основаниям соответствуют вертикальная и горизонтальная координаты; третье основание показано по вертикали справа, оно делает двумерную таблицу трехмерным кубом 4 х 4 х 4.
Физических промежутков между кодонами нет, поскольку код характеризуется непрерывностью. Если бы код был синглетным , то есть если бы каждой кодируемой аминокислоте соответствовало бы только одно основание (из четырех), кодирующая емкость кода и составляла бы только четыре аминокислоты. Между тем, таких аминокислот двадцать, и только этого числа (не меньше!) достаточно для обеспечения существующего белкового разнообразия. Если бы код был дублетным , то есть, если бы каждой кодируемой аминокислоте соответствовало бы два основания, кодирующая емкость кода составляла бы только шестнадцать аминокислот (42) – то есть очевидно недостаточно. Емкость триплетного кода составляет 64 аминокислоты (43). Этого хватает с избытком.
«Избыток» составляет 44 кодируемых продукта. Многие исследователи утверждают, что эволюция генетического кода шла в направлении от синглетного к триплетному. При этом они не понимают, что смена размера кодирующей единицы потребовала бы принципиального изменения всей машины кодирования (то есть всего набора ферментов, обслуживающих этот процесс) – вещь невозможная! Поэтому синглетный этап триплетного кодирования (если он был) мог означать, что в составе триплета значащей единицей могла быть только одна (например, первая или любая), а в составе дублета – две. Тогда и непрерывность кодированной записи могла быть только физической. Функционально значащие основания разделялись остальными основаниями триплета, а эволюция продолжала совершенствовать структуру молекул-участников кодирования. Произвольные (из четырех) третьи основания современных кодонов для восьми (из двадцати) аминокислот могут быть реликтами до-триплетных кодов. Выбор же тройки азотистых оснований в качестве дискретной единицы генетического кода мог быть обусловлен также термодинамикой взаимодействия пар оснований, при котором матричное копирование инициируется их триплетом (мы говорили об этом выше, ссылаясь на Зенгера).
Неперекрываемость – один и тот же нуклеотид не может входить одновременно в состав двух или более триплетов; не соблюдается для некоторых перекрывающихся генов вирусов, митохондрий и бактерий, которые кодируют несколько белков, считывающихся со сдвигом рамки.
Поскольку никакого избытка в крайне экономной природе не бывает, он и здесь компенсируется еще одним свойством, парадоксально именуемым вырожденностью( избыточностью), которая означает, что каждую аминокислоту (два исключения – метионин и триптофан) кодирует более, чем один триплет. Синим выделены клетки таблицы, содержащие аминокислоту, кодируемую четырьмя триплетами, серым – тремя, светло-серым – двумя, самым светлым – две аминокислоты, кодируемые только одним триплетом.
Читать дальшеИнтервал:
Закладка:








