Е. Бессолицына - Структурная биохимия

- Название:Структурная биохимия
- Автор:
- Жанр:
- Издательство:Array Литагент «Ридеро»
- Год:неизвестен
- ISBN:9785447442668
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Е. Бессолицына - Структурная биохимия краткое содержание
Структурная биохимия - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Структурные полисахариды, входящие в состав межклеточного вещества, образуют протеогликаны – белково-углеводные комплексы, где белок составляет около 10%. Типичный протеогликан хрящевой ткани содержит около 150 полисахаридных цепей с молекулярной массой 20000 каждая; они (в виде боковых цепей) ковалентно присоединены к «сердцевинным» полипептидам. Такие протеогликаны представляют собой сильно гидратированные структуры, 20000 каждая; они (в виде боковых цепей) ковалентно присоединены к «сердцевинным» полипептидам. Такие протеогликаны представляют собой сильно гидратированные структуры.
Гликопротеиды – это молекулы белка, к которым присоединены короткие полисахаридные молекулы. В отличие от протеогликанов, в гликопротеидах белок составляет 95% молекулы. Кроме того, углеводный компонент короче – несколько десятков мономерных звеньев, очень разнообразен, можно сказать, что каждый белок имеет свой уникальный как по составу, и по форме полисахарид. Полисахариды присоединяются к секретируемым белкам клетки, а также к мембранным белкам, углеводная часть протеидов мембраны обращена во внешнюю среду клетки, формирует гликокаликс.
В данном случае полисахарид выполняет сигнальную функцию, по таким разветвленным гетерополисахаридам клетки распознают друг друга. В частности группы крови А В определяются по полисахаридам располагающимся на поверхности эритроцитов. Здесь можно говорить не только о многообразии полисахаридного компонента каждого белка, но и о вариабельности этой молекулы для одного белка внутри популяции.
Нуклеотиды и нуклеиновые кислоты
Структура нуклеотидов и азотистых оснований
Нуклеотиды принимают участие во множестве биохимических процессов, а также являются мономерами нуклеиновых кислот. Нуклеиновые кислоты обеспечивают все генетические процессы. Каждый нуклеотид состоит из трех типов химических молекул:
• азотистое основание;
• моносахарид;
• 1—3 остатка фосфорной кислоты.
В отличие от моносахаридов, нуклеотиды как мономеры являются сложно устроенными молекулами, состоящими из структур, относящихся к разным классам химических веществ, поэтому необходимо рассмотреть свойства и структуру этих компонентов по отдельности.
Азотистые основания
Азотистые основания относятся к гетероциклическим соединениям. В состав гетероцикла помимо атомов углерода входят атомы азота. Все азотистые основания, входящие в нуклеотиды относят к двум классам азотистых оснований: пуриновые и пиримидиновые. Пуриновые основания это производные пурина – гетероцикла, состоящего из двух циклов, один пятичленный, второй – шести, нумерация осуществляется так, как показано на рисунке. Пиримидиновые основания являются производными пиримидина и состоят из одного шестичленного цикла, нумерация также указана на рисунке (Рисунок 31). Главные пиримидиновые основания и у прокариот, и у эукариот – это цитозин, тимини урацил.Из пуриновых оснований чаще всего встречаются аденини гуанин.Два других – ксантини гипоксантин – являются интермедиатами в процессах их метаболизма. У человека в роли конечного продукта катаболизма пуринов выступает окисленное пуриновое основание – мочевая кислота. Помимо пяти названных выше главных оснований известны и менее широко представленные минорные основания. Некоторые из них присутствуют только в нуклеиновых кислотах бактерий и вирусов, но многие также найдены в составе про- и эукариотических ДНК и транспортных и рибосомных РНК. Так, и бактериальная ДНК, и ДНК человека содержат значительные количества 5-метилцитозина; в бактериофагах обнаружен 5-гидроксиметилцитозин. Необычные основания выявлены в матричной РНК – N 6-метиладенин, N 6, N 6-диметиладенин и N 7-Meтилгуанин. У бактерий также обнаружен модифицированный урацил с присоединенной по N 3-положению (α-амино, α-карбокси) -пропильной группой. Функции этих замещенных пуринов и пиримидинов до конца не выяснены, однако они могут образовывать неканонические связи между основаниями (это будет рассмотрено ниже), обеспечивая образование вторичных и третичных структур нуклеиновых кислот.
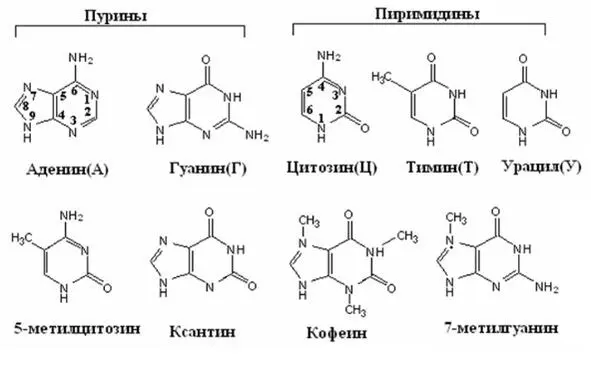
Рисунок 31. Структура азотистых оснований
В клетках растений выявлена серия пуриновых оснований с метильными заместителями. Многие из них фармакологически активны. В качестве примера можно привести кофейные зерна, содержащие кофеин (1,3, 7-триметилксантин), чайный лист, содержащий теофиллин (1, 3-диметил-ксантин), и какао-бобы, в состав которых входит теобромин (3, 7-диметилксантин).
изомерия и Физико-химические свойства пуриновых и пиримидиновых оснований
Молекула азотистого основания образует систему чередующихся одинарных и двойных связей (систему сопряженных двойных связей). Такая организация образует жесткую молекулу, без возможности конформационных переходов. В результате нельзя говорить об изменении конформации азотистых оснований.
Для азотистых оснований выявлен только один тип изомерии кето-енольный переход или таутомерия.
Таутомерия
Благодаря феномену кето-енольной таутомерии нуклеотиды могут существовать либо в лактимной, либо в лактамной формах, причем в физиологических условиях лактамная форма превалирует у гуанина и тимина (Рисунок 32). Важность этого обстоятельства станет ясна при обсуждении процессов спаривания оснований.
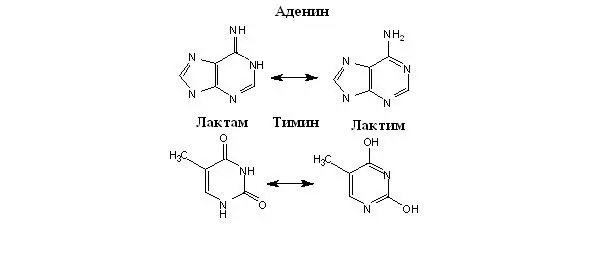
Рисунок 32. Таутомерия нуклеотидов
Растворимость
При нейтральном рН наименьшей растворимостью обладает гуанин. Следующим в этом ряду стоит ксантин. Мочевая кислота в форме уратов сравнительно неплохо растворяется при нейтральном рН, но очень плохо растворима в жидкостях с более низкими значениями рН, таких, как моча. Гуанин в моче человека в норме отсутствует, а ксантин и мочевая кислота являются ее обычными компонентами. Последние два пурина часто входят в состав камней мочевого тракта.
Поглощение света
За счет системы сопряженных двойных связей все азотистые основания поглощают в ультрафиолетовой части спектра. Спектр поглощения – график распределения оптической плотности в зависимости от длины волны. Для каждого азотистого основания свой спектр поглощения, по нему можно различить растворы различных азотистых оснований или соединений в состав которых входит азотистое основание (нуклеотиды), но максимум поглощения у всех совпадает при длине волны 260 нм. Это позволяет легко и быстро определять концентрацию как азотистых оснований, так нуклеотидов и нуклеиновых кислот. Спектр поглощения также зависит от рН раствора (Рисунок 33).
Читать дальшеИнтервал:
Закладка:









