Ричард Докинз - Слепой часовщик

- Название:Слепой часовщик
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Ричард Докинз - Слепой часовщик краткое содержание
Книга посвящена современным аспектам теории эволюции. Докинз — убежденный дарвинист, и в свойственном ему легком стиле, но без всяких упрощений, излагает наиболее сложные положения теории Дарвина настолько ясно, что в верности этой теории не остается никаких сомнений.
The Blind Watchmaker by Richard Dawkins. Перевод с английского: Анатолий Протопопов.
Слепой часовщик - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Здесь читатель может быть озадачен явной непоследовательностью. Вся эта книга подчеркивает первостепенную важность естественного отбора. Как же мы теперь можем подчеркивать хаотичность эволюционных изменений на молекулярном уровне? Предвосхищая главу 11, скажу, что на деле здесь нет никакой вражды с эволюцией адаптаций, что является главной темой этой книги. Даже самый горячий нейтралист не думает, что сложные рабочие органы, подобные глазам и рукам развились посредством случайного дрейфа. Всякий нормальный биолог согласится, что они могут развиться лишь посредством естественного отбора. Нейтралисты лишь полагают (и по моему мнению — справедливо), что такие адаптации — лишь вершина айсберга: вероятно, наибольшее количество эволюционных изменений, а именно — изменений на молекулярном уровне, бесфункционально.
Поскольку молекулярные часы — это факт, и похоже на правду, что молекулы определённого вида изменяются за миллионы лет с присущим им характерным темпом, то мы можем использовать этот факт для датирования точек разветвления эволюционного древа. И если действительно верно, что большая часть эволюционных изменений на молекулярном уровне нейтральна, то это замечательный подарок для систематика. Это означает, что проблема конвергенции может быть снята при помощи статистики. Каждое животное имеет большой объём генетического текста, хранящегося в его клетках, и большая часть его, согласно нейтралистской теории, не имеет никакого отношения к его приспособленности к данным условиям жизни; текст, который в значительной степени не затрагивается отбором и в значительной степени не является объектом конвергентной эволюции, кроме как в результате явной случайности. Вероятность того, что два больших фрагмента селективно-нейтрального текста будут случайно походить друг на друга, можно рассчитать, и она очень и очень низка. Еще важнее, что постоянный темп молекулярной эволюции позволяет нам практически датировать точки разветвления эволюционной истории.
Трудно преувеличить ту мощь, которую новые методы чтения молекулярных последовательностей добавили к арсеналу систематика. Конечно, пока ещё не все молекулярные последовательности, и не всех животных, расшифрованы, но уже можно пойти в библиотеку и посмотреть точную, дословную и побуквенную последовательность молекулярного предложения, такого как, скажем, «гемоглобин» в варианте собаки, кенгуру, колючего муравьеда, цыплёнка, гадюки, тритона, карпа и человека. Не у всех животных есть гемоглобин, но есть другие белки, например гистоны, своя версия которых существует в каждом животном и растении, и опять же — многие из них можно уже посмотреть в библиотеке. И это не зыбкие характеристики, могущие, подобно длине ног или ширине черепа, зависеть от возраста и здоровья экземпляра, или даже от особенностей зрения исследователя. Это точно сформулированные альтернативные версии одного и того же предложения, на одном и том же языке, которые могут быть положены рядом и сравнены так же скрупулёзно и точно, как скрупулёзно греческий учёный мог бы сравнивать два пергамента с текстом одного и того же Евангелия. ДНК-последовательности — Евангелия всей жизни, и мы научились их расшифровывать.
Базовое положение систематики гласит, что близкие родственники будут иметь более совпадающие версии конкретного молекулярного предложения, чем более отдалённые. Это называется «принцип экономии». Экономия — другое название экономической скупости. Наш задача состоит в том, что исходя из набора животных, предложения которых известны, скажем, восьми животных, перечисленных в предыдущем абзаце, выяснить, какая из всех возможных древовидных диаграмм соединяет эти восемь животных наиболее экономно. Самое экономное дерево — это «экономически скупейшее» дерево предположений, в том смысле, что оно предполагает минимальное количество изменений слов в эволюции и минимальное количество конвергенций. Мы вправе предполагать минимальное количество конвергенций на основании явного неправдоподобия. Маловероятно, особенно если учесть, что большая часть молекулярной эволюции нейтральна, что два неродственных животных натолкнутся на в точности ту же самую последовательность, дословно и побуквенно.
При попытке рассмотреть на все возможные деревья обнаруживаются вычислительные трудности. Когда нам нужно расклассифицировать только трёх животных, то возможных деревьев будет только три: А вместе с B, исключая C; А с C, исключая B; и B с C, исключая A. Можно проделать аналогичные вычисления для больших количеств животных, подлежащих классификации, но количество возможных деревьев круто возрастает. Когда нам нужно рассмотреть только четверых животных, то общее количество возможных деревьев родства равно 15 и пока что обозримо. Компьютеру не потребуется долго работать, чтобы выяснить, какое из этих 15 является наиболее скупым. Но если нужно рассмотреть 20 животных, то число возможных деревьев, составит 8 200 794 532 637 891 559 375 (см. рисунок 9). Наибыстрейшему на сегодняшний день компьютеру потребуется 10 000 миллионов лет (примерно возраст Вселенной), чтобы вычислить скупейшее дерево для всего 20 животных. А систематики часто хотят строить деревья из более чем 20 животных.
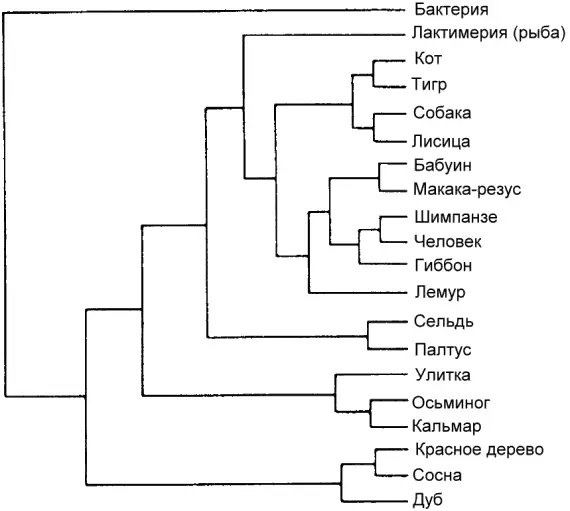
Рис. 9.
Это генеалогическое древо правильно. Существуют также 8200794532637891559374 других способа классификации этих 20 организмов, и все они неправильны.
Хотя молекулярные систематики были первыми, осознавшие эту проблему взрывного роста вариантов, фактически она уже существовала и в недрах немолекулярной таксономии. Немолекулярные систематики просто уклонились от неё, делая интуитивные предположения. Из всех, в принципе возможных генеалогических деревьев, подавляющее большинство может быть отброшено сразу — например, все миллионы генеалогических деревьев, которые располагают людей ближе к дождевым червям, чем к шимпанзе. Систематики даже не взглянут на такие очевидно абсурдные деревья родства, но сосредоточатся на относительно немногих деревьях, которые не слишком радикально нарушают их предположения. Это, видимо, справедливо, хотя всегда есть опасность, что правильное, скупейшее дерево оказалось одним из тех, что были отвергнуты без рассмотрения. Компьютеры также можно запрограммировать на использование кратчайших путей, тем самым проблему взрывообразно больших чисел можно милосердно сократить.
Молекулярная информация настолько обильна, что позволяет нам многократно и независимо строить системы родства на основе различных белков. При этом мы можем использовать выводы, сделанные при изучении одной молекулы, для проверки выводов, сделанных на основании другой. Если нас беспокоит, что история, рассказанная одной белковой молекулой, на деле может быть результатом конвергенции, то мы можем безотлагательно проверить её, изучив другую. Конвергентная эволюция — это собственно особый вид случайного стечения обстоятельств. Особенность случайных совпадений состоит в том, что, даже если они, несмотря на редкость, однажды всё-таки случаются, то просто повторение их вероятно уже гораздо менее. И уж тем более низка их вероятность при трёхкратном повторении. Рассматривая всё больше и больше независимых белковых молекул, мы можем практически исключить случайное совпадение.
Читать дальшеИнтервал:
Закладка:





