Александр Солодков - Физиология человека. Общая. Спортивная. Возрастная

- Название:Физиология человека. Общая. Спортивная. Возрастная
- Автор:
- Жанр:
- Издательство:Литагент Спорт
- Год:2016
- Город:Москва
- ISBN:978-5-906839-67-1
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Александр Солодков - Физиология человека. Общая. Спортивная. Возрастная краткое содержание
Для студентов, аспирантов, научных сотрудников, преподавателей, тренеров и врачей, работающих в области физической культуры.
Физиология человека. Общая. Спортивная. Возрастная - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Отдавший кислород гемоглобин считают восстановленным, или дезоксигемоглобином. Поскольку молекула гемоглобина содержит 4 частицы гема (железосодержащего вещества), она может связать четыре молекулы О 2. Количество О 2, связанного гемоглобином в 100 мл крови, носит название кислородной емкости крови и составляет около 20 мл О 2. Кислородная емкость всей крови человека, содержащей примерно 750 г гемоглобина, приблизительно равна 1 л.
Каждому значению рО 2в крови соответствует определенное процентное насыщение гемоглобина кислородом. Кривую зависимости процентного насыщения гемоглобина кислородом от величины парциального напряжения называют кривой диссоциации оксигемоглобина (рис. 21). Анализ хода этой кривой сверху вниз показывает, что с уменьшением рО 2в крови происходит диссоциация оксигемоглобина, т. е. процентное содержание оксигемоглобина уменьшается, а восстановленного – растет.
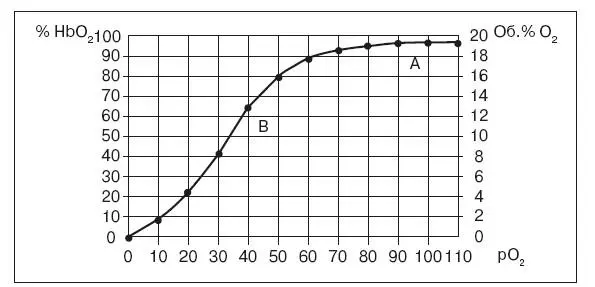
Рис. 21. Кривая диссонации оксигемоглобина в крови человека в покое:
А – содержание HbО 2в артериальной крови. В – то же в венозной крови
В различных условиях деятельности может возникать острое снижение насыщенности крови кислородом– гипоксемия. Причины гипоксемии весьма разнообразны. Она может развиваться вследствие снижения рО 2в альвеолярном воздухе (произвольная задержка дыхания, вдыхание воздуха с пониженным рО 2), при физических нагрузках, а также при неравномерной вентиляции различных отделов легких.
Образующийся в тканях СО 2диффундирует в тканевые капилляры, откуда переносится венозной кровью влегкие, где переходит в альвеолы и удаляется с выдыхаемым воздухом. Углекислый газ в крови (как и О 2) находится в двух состояниях: растворенный в плазме (около 5 % всего количества) и химически связанный с другими веществами (95 %). СО, в виде химических соединений имеет три формы: угольная кислота (Н 9СО 3), соли угольной кислоты (NaHCО 3) и в связи с гемоглобином (HbНСО 3).
В крови тканевых капилляров одновременно с поступлением СО 2внутрь эритроцитов и образованием в них угольной кислоты происходит отдача О 2оксигемоглобином. Восстановленный Hb легко связывает водородные ионы, образующиеся при диссоциации угольной кислоты. Таким образом, восстановленный Hb венозной крови способствует связыванию СО 2, а оксигемоглобин, образующийся в легочных капиллярах, облегчает его отдачу.
В состоянии покоя с дыханием из организма человека удаляется 230–250 мл СО 2в 1 минуту. При удалении из крови СО 2из нее уходит примерно эквивалентное число ионов водорода. Таким порядком дыхание участвует в регуляции кислотно-щелочного состояния во внутренней среде организма.
Обмен газов между кровью и тканями осуществляется также путем диффузии. Между кровью в капиллярах и межтканевой жидкостью существует градиент напряжения О 2, который составляет 30–80 мм рт. ст., а напряжение СО 2в интерстициальной жидкости на 20–40 мм рт. ст. выше, чем в крови. Кроме того, на обмен О 2и СО 2в тканях влияют площадь обменной поверхности, количество эритроцитов в крови, скорость кровотока, коэффициенты диффузии газов в тех средах, через которые осуществляется их перенос.
Артериальная кровь отдает тканям не весь О 2. Разность между об.% О 2в притекающей к тканям артериальной крови (около 20 об.%) и оттекающей от них венозной кровью (примерно 13 об.%) называется артерио-венозной разностью по кислороду (7 об.%). Эта величина служит важной характеристикой дыхательной функции крови, показывая, какое количество О, доставляют тканям каждые 100 мл крови. Для того чтобы установить, какая часть приносимого кровью О 2переходит в ткани, вычисляют коэффициент утилизации (использования) кислорода. Его определяют путем деления величины артериовенозной разности на содержание О 2в артериальной крови и умножения на 100. В покое для всего организма коэффициент утилизации О 2равен примерно 30–40 %. Однако в миокарде, сером веществе мозга, печени и корковом слое почек он составляет 40–60 %. При тяжелых физических нагрузках коэффициент утилизации кислорода работающими скелетными мышцами и миокардом достигает 80–90 %.
В снабжении мышц О 2при тяжелой работе имеет определенное значение внутримышечный пигмент миоглобин, который связывает дополнительно 1,0–1,5 л О 2. Связь О 2с миоглобином более прочная, чем с гемоглобином. Оксимиоглобин отдает О 2только при выраженной гипокеемии.
10.3. Регуляция дыхания
Регуляция внешнего дыхания представляет собой физиологический процесс управления легочной вентиляцией для обеспечения оптимального газового состава внутренней среды организма в постоянно меняющихся условиях его жизнедеятельности. Основную роль в регуляции дыхания играют рефлекторные реакции, возникающие в результате возбуждения специфических рецепторов, заложенных в легочной ткани, сосудистых рефлексогенных зонах и скелетных мышцах. Центральный аппарат регуляции дыхания представляют нервные образования спинного, продолговатого мозга и вышележащих сегментов ЦНС.
Гуморальная регуляция дыхания, созданная Д. Холденом и Д. Пристли около 50 лет назад, в последние годы не находит экспериментального подтверждения, большинством специалистов считается ошибочной и упоминается сейчас только в историческом плане. Это обусловлено открытием специфических рецепторов (механо- и хеморецепторов), а также других рефлекторных влияний на дыхательный центр. Поэтому все изменения внешнего дыхания в настоящее время объясняются только рефлекторными механизмами.
Дыхательный ритм и управление деятельностью дыхательных мышц генерируются работой дыхательного центра, представляющего собой совокупность взаимосвязанных нейронов ретикулярной формации продолговатого мозга и вышележащих отделов ЦНС, обеспечивающих тонкое приспособление дыхания к различным условиям внешней среды. Современные представления о работе дыхательного центра сводятся к тому, что часть дыхательных нейронов, объединенных в так называемую латеральную зону, является эфферентной частью дыхательного центра и обеспечивает преимущественно фазу вдоха (инспиратор – ные нейроны). Другая группа нейронов, составляющая медиальную зону, является афферентной частью дыхательного центра и обеспечивает фазу выдоха (экспираторные нейроны). Предназначение этой зоны заключается в контроле за периодичностью дыхательной ритмики, организуемой латеральной зоной.
Читать дальшеИнтервал:
Закладка: