Коллектив Авторов - Цифровой журнал «Компьютерра» № 108

- Название:Цифровой журнал «Компьютерра» № 108
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Коллектив Авторов - Цифровой журнал «Компьютерра» № 108 краткое содержание
Кафедра Ваннаха: Горячая цифровая война Автор: Михаил Ваннах
Дмитрий Шабанов: Парадокс ускорения эволюции Автор: Дмитрий Шабанов
Кивино гнездо: Пароль для серой зоны Автор: Киви Берд
Василий Щепетнёв: Предсказания и пророчества Автор: Василий Щепетнев
Дмитрий Вибе: Мы сверху Автор: Дмитрий Вибе
Голубятня-ОнлайнГолубятня: Идеальный мобильный звук. Часть вторая Автор: Сергей Голубицкий
Цифровой журнал «Компьютерра» № 108 - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Но любой ли признак зависит от одного гена? Нет. К примеру, в только что упомянутом эксперименте по эволюции термоустойчивости бактерий эффект многих мутаций невозможно было оценить по отдельности, без учета аллельного состояния других генов. Одна и та же мутация в комплекте с одними аллелями иных генов оказывается благоприятной, в сочетании с другими — вредной. Это значит, что определяющее судьбу состояние признака зависит от нескольких генов.
Снижают ли взаимодействия между генами эффективность отбора? Да, и особенно для видов с половым размножением. Я приведу пример именно для таких организмов, каждый из которых имеет по два одинаковых или разных аллеля каждого гена и передаёт потомкам лишь один из них.
Представим, что некий счастливый организм приобретает сочетание двух редких (возможно, недавно возникших) аллелей, каждый из которых поодиночке неблагоприятен: А 1и В 1. В популяции распространены аллели А 0и В 0. Сочетания А 1и В 1благоприятны, а вот А 1и В 0, как и А 0и В 1— вредны (и то и другое — в сравнении с нормой, А 0и В 0).
Давайте я не буду объяснять логику решения генетических задач, а просто распишу соотношения численностей групп потомков от скрещивания счастливого обладателя удачного сочетания двух мутаций с наиболее вероятным партнёром. Рассмотрим случаи, когда оба этих аллеля доминантны (влияют на фенотип даже у особей, у которых присутствуют в одной копии) и когда они рецессивны (проявляются в фенотипе только у организма, имеющего две их копии).
Если аллели А 1и В 1доминантны, только четверть потомков будет обладать улучшенным фенотипом (показаны полужирным шрифтом), а две четверти — ухудшенным (выделены подчёркиванием):
А А 1А 0В 1В 0 × А 0А 0В 0В 0 → А 1А 0В 1В 0 : 1А 0В 0В 0 0А 0В 1В 0 0А 0В 0В 0.
Если рассматриваемые аллели рецессивны, у всех потомков будет нормальный фенотип:
А 1А 1В 1В 1 × А 0А 0В 0В 0 → А 1А 0В 1В 0.
Посмотрим, что будет, если такие потомки скрестятся друг с другом:
А 1А 0В 1В 0 × А 1А 0В 1В 0 → А 1А 1В 1В 1: 2 0А 1В 1В 1 1А 1В 1В 0 1А 0В 1В 0: 2А 1А 0В 0В 0: 2А 0А 0В 1В 0: 0А 0В 1В 1 1А 1В 0В 0 0А 0В 0В 0.
Благоприятный признак проявится лишь у одного потомка из шестнадцати, а шесть из шестнадцати потомков будут иметь ухудшенный фенотип! В этих (и иных, не рассмотренных нами) случаях подобного взаимодействия аллелей они чаще будут встречаться в популяции порознь, когда отбор будет работать против них, чем вместе, в одобряемом отбором состоянии. Ситуации, когда отбор будет поддерживать взаимодополняющую пару аллелей, окажутся редки по сравнению со случаями, когда он будет уничтожать эти аллели поодиночке.
Взаимодействие генов снижает эффективность отбора! И чем сложнее организмы, чем сложнее определяются их признаки, тем в меньшей мере направленный отбор фенотипов будет приводить к направленному отбору аллелей.
У по-настоящему сложных организмов между генотипом и фенотипом вклинивается ещё несколько механизмов регуляции, затрудняющих сортировку аллелей отбором. Один из них на языке СТЭ называется расширением нормы реакции. Организмы с одинаковым генотипом могут отличаться из-за развития в разных условиях (или просто в силу случайности). Представьте себе трёх человеческих близнецов-тройняшек с одинаковыми генотипами. Одного вырастило племя первобытных охотников, другой попал в интернат для спортсменов-силовиков, третий — в спецматшколу. Несмотря на идентичную наследственность, они вырастут очень разными — и физически, и по поведению. В отличие от ненаправленных мутаций, их отличия повысят приспособленность их носителей. При отборе (например, половом — покорении сердца потенциально плодовитой красавицы) эти три близнеца будут иметь разные шансы, ведь отбор (и половой в том числе) оценивает именно фенотипы!
Второй механизм связан с эпигенетической регуляцией генной активности, особенно характерной для млекопитающих. В зависимости от состояния организма, он передаёт потомкам геномы с разными химическими «метками» на генах (метилированием или иными изменениями «букв» генетического текста). Эти метки влияют на активность генов и, значит, на признаки. Обычно эпигенетические метки обеспечивают тот вариант развития (в пределах нормы реакции), который более соответствует данным условиям среды. При изменившихся условиях эти метки могут быть стёрты или переставлены.
Как эти изменения влияют на эффективность отбора? Новую схему со многими поколениями рисовать не буду, покажу лишь логику формирования признаков одной особи.
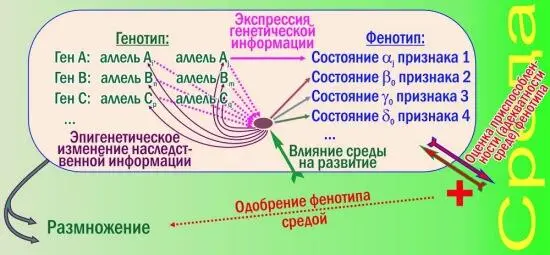
Кроме основной петли обратной связи (генотип влияет на фенотип; отбирая лучшие фенотипы, среда отбирает лучшие генотипы) появились новые уровни регуляции. Смотрите: развитие признака 1 по-прежнему управляется достаточно просто, он зависит от одного аллеля. Зато признаки 2, 3 и 4 зависят и от всех показанных генов, и от среды, причём в разной степени. Взаимодействие генных продуктов и средовых влияний формирует сложнейший узел взаимопереплетённых причин и следствий. От результатов такого взаимодействия зависит и сама генная активность: в зависимости от него гены подвергаются эпигенетическому редактированию — обратимому наследственному изменению.
Эти изменения снижают эффективность сортировки аллелей отбором. Связь фенотипов с генотипами становится очень неоднозначной (кроме «простых» признаков, как признак 1 на рис. 2). Механизм эволюции, эффективно работавший на рис. 1, будет пробуксовывать на рис. 2.
Эволюция — это соревнование в скорости выработки новых приспособлений. Вернитесь к первому рисунку: во втором показанном там поколении выигрывал нормальный генотип, а в третьем носитель недавней нормы уже проиграл, так как появились более приспособленные организмы. Как же могли возникнуть организмы, у которых эффективность отбора (и эффективность выработки новых приспособлений) понизилась, — они ведь неминуемо должны были проигрывать тем, кто эволюционировал быстрее?
Дело в том, что у сложных организмов эффективность отбора не снижается, а вырастает. Опять процитирую Расницына:
"...возраст (в миллионах лет) полусовременной фауны (в которой половина видов — современные, половина — вымершие) у крупных млекопитающих (хоботные и копытные) — 0,2, у мелких — 0,5, у птиц и рыб — 0,7, у насекомых — 3-7, у моллюсков — 3,5-5, у диатомовых водорослей — 15. Время полувымирания (по аналогии с полураспадом, время вымирания половины исходного числа видов) в тех же единицах наименьшее у слонов — 0,18, а в среднем у млекопитающих — 0,54, у костистых рыб — 3,5, у граптолитов — 1,3, иглокожих — 4,2, двустворок — 7, у планктонных фораминифер — 5, бентосных — 18-24, у диатомовых водорослей — 5,5, у динофлагеллат — 9. На старших таксономических уровнях различия подобные, но часто более резкие. Например, возраст полусовременной фауны для родов млекопитающих — 4 млн лет, для птиц — 10, рептилий — 20, рыб — 30-50, насекомых — 40, моллюсков — 60, фораминифер — 230".
Читать дальшеИнтервал:
Закладка: