БСЭ БСЭ - Большая Советская Энциклопедия (БИ)

- Название:Большая Советская Энциклопедия (БИ)
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
БСЭ БСЭ - Большая Советская Энциклопедия (БИ) краткое содержание
Большая Советская Энциклопедия (БИ) - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Пространственная структура нативной ДНК образована двумя комплементарными нитями и представляет собой двойную спираль Крика — Уотсона; в ней противоположные азотистые основания попарно связаны водородными связями — аденин с тимином и гуанин с цитозином. Устойчивость двойной спирали обеспечивается, наряду с водородными связями, также гидрофобным взаимодействием между плоскими кольцами азотистых оснований, расположенных стопкой (стопочное взаимодействие, или стакинг). Нити РНК спирализованы лишь частично. ДНК вирусов, бактериофагов, бактерий, а также митохондриальная в ряде случаев представляет собой замкнутое кольцо; при этом наряду со спиралью Крика — Уотсона наблюдается ещё дополнительная т. н. сверхспирализация.
Денатурация Б. Нарушение нативной пространственной структуры Б. при различных воздействиях (повышение температуры, изменение концентрации металлов, кислотности раствора и др.) называется денатурацией и в ряде случаев обратимо (обратный процесс называется ренатурацией; рис. 2 ). Молекулы Б. — кооперативные системы: поведение их зависит от взаимодействий составляющих частей. Кооперативность молекул Б. определяется тем, что повороты отдельных звеньев из-за внутримолекулярных взаимодействии зависят от конформации соседних звеньев. В основе денатурации Б. при изменении внешних условий обычно лежат кооперативные конформационные превращения (например, переходы a-спираль — b-структура, a-спираль — клубок, b-структура — клубок для полипептидов, переход глобула — клубок для глобулярных белков, переход спираль — клубок для нуклеиновых кислот). В отличие от фазовых переходов (кипение жидкости, плавление кристалла), являющихся предельным случаем кооперативных процессов и происходящих скачком, кооперативные переходы Б. совершаются в конечном, хотя и сравнительно узком, интервале изменений внешних условий. В этом интервале одномерные, линейные молекулы (нуклеиновые кислоты, полипептиды), претерпевающие переход спираль — клубок, разбиваются на чередующиеся спиральные и клубкообразные участки ( рис. 3 ).
Переход спираль — клубок в ДНК наблюдается при повышении температуры, добавлении в раствор кислоты или щёлочи, а также под влиянием других денатурирующих агентов. Этот переход в гомополинуклеотидах происходит при нагревании в интервале десятых долей °С, в фаговых и бактериальных ДНК — в интервале 3—5°С ( рис. 3 ), в ДНК высших организмов — в интервале 10—15 °С. Чем выше гетерогенность ДНК, тем шире интервал перехода и меньше способность молекул ДНК к ренатурации. Переход спираль — клубок в различных видах РНК носит менее кооперативный характер ( рис. 4 ) и происходит в более широком интервале температурных или других денатурирующих воздействий.
Б. — полимерные электролиты, их пространственная конформация и кооперативные переходы зависят как от степени ионизации молекулы, так и от концентрации ионов в среде, что влияет на электростатические взаимодействия как между отдельными частями молекулы, так и между Б. и растворителем.
Строение и биологические функции Б. Строение Б. — результат длительной эволюции на молекулярном уровне, вследствие чего эти молекулы идеально приспособлены к выполнению своих биологических задач. Между первичной структурой, конформацией Б. и конформационными переходами, с одной стороны, и их биологическими функциями — с другой, существуют тесные связи, исследование которых — одна из главных задач молекулярной биологии . Установление таких связей в ДНК позволило понять основные механизмы репликации , транскрипции и трансляции , а также мутагенеза и некоторых других важнейших биологических процессов. Линейная структура молекулы ДНК обеспечивает запись генетической информации, её удвоение при матричном синтезе ДНК и получение (также путём матричного синтеза) многих копий с одного и того же гена, т. е. молекул и-РНК. Сильные ковалентные связи между нуклеотидами обеспечивают сохранность генетической информации при всех этих процессах. В то же время относительно слабые связи между нитями ДНК и возможность вращения вокруг простых химических связей обеспечивают гибкость и лабильность пространственной структуры, необходимые для разделения нитей при репликации и транскрипции, а также подвижность молекулы и-РНК, служащей матрицей при биосинтезе белка (трансляция). Исследование пространственной структуры и конформационных изменений белков-ферментов на разных стадиях ферментативной реакции при взаимодействии с субстратами и коферментами даёт возможность установить механизмы биокатализа и понять природу огромного ускорения химических реакций, осуществляемого ферментами.
Методы исследования Б. При исследовании строения и конформационных превращений Б. широко используются как очищенные природные Б., так и их синтетические модели, которые проще по строению и легче поддаются исследованию. Так, при изучении белков моделями служат гомогенные или гетерогенные полипептиды (с заданным или случайным чередованием аминокислотных остатков). Моделями ДНК и РНК являются соответствующие синтетические гомогенные или гетерогенные полинуклеотиды. К методам исследования Б. и их моделей относятся рентгеноструктурный анализ, электронная микроскопия, изучение спектров поглощения, оптической активности, люминесценции, методы светорассеяния и динамического двойного лучепреломления, седиментационный метод, вискозиметрия, физико-химические методы разделения и очистки и ряд др. Все методы, разработанные для изучения синтетических полимеров, применимы и к Б. При трактовке свойств Б. и их моделей, закономерностей их конформационных превращений используются также методы теоретической физики (статистической физики, термодинамики, квантовой механики и др.).
Лит.: Бреслер С. Е., Введение в молекулярную биологию, М.—Л., 1966; Волькенштейн М. В., Молекулы и жизнь, М., 1965; Уотсон Дж., Молекулярная биология гена, пер. с англ., М., 1967; физические методы исследования белков и нуклеиновых кислот, М., 1967.
Ю. С. Лазуркин.
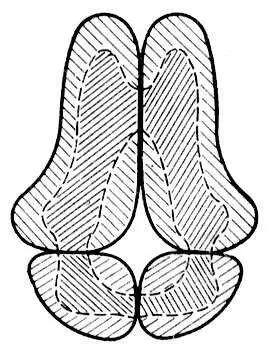
Рис. 1. Образование четвертичной структуры глобулярных белков. Заштрихованы редко — полярные (гидрофильные) части белковых глобул, густо — неполярные (гидрофобные) области.
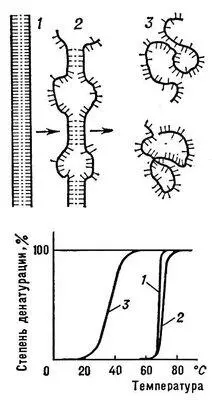
Рис. 3. Схема перехода спираль — клубок для ДНК: 1 — нативное состояние (вместо двойной спирали для простоты изображена «верёвочная лестница»); 2 — состояние ДНК в области перехода; 3 — денатурированное состояние (однонитевые клубки).
Читать дальшеИнтервал:
Закладка: