Кирилл Еськов - Доказательства эволюции

- Название:Доказательства эволюции
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:2010
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Кирилл Еськов - Доказательства эволюции краткое содержание
). Отличия файла от оригинала минимальны. В файле отсутствуют по понятным причинам несколько видеороликов, которые имеются в оригинале, но все изображения и гиперссылки сохранены. Кроме того, в файле отсутствует самая последняя глава — фрагменты из книги Ф. Коллинза.
Доказательства эволюции - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Быстрое развитие изоляции у лососевых рыб.Быстрое возникновение репродуктивной изоляции при расхождении по «экологическим» параметрам характерно отнюдь не только для членистоногих, о которых шла речь выше. Так, показано возникновение частичной репродуктивной изоляции между двумя популяциями нерки ( Oncorhynchus nerka ), которые начали расходиться от одной предковой линии не ранее, чем в 1937 г. В указанном году нерка была интродуцирована в оз. Вашингтон (штат Вашингтон, США) и вскоре образовала крупную местную популяцию во впадающей в озеро реке (Cedar River). В 1957 г. было обнаружено, что часть особей стала нереститься не в реке, а в озере. В 1992 г. было установлено, что «речные» и «озерные» формы уже представляют собой две частично изолированные популяции, немного отличающиеся морфологически. Некоторые представители «речной» популяции иногда нерестятся в озере, в тех же местах, что и «озерные» формы, однако даже в этом случае скрещивания между представителями разных популяций происходят, по-видимому, значительно реже, чем это диктуется простой вероятностью (Hendry et al., 2000). Репродуктивная изоляция в данном случае развилась самое большее за 56 лет, что соответствует примерно 13 поколениям.
Все эти факты говорят о том, что репродуктивная изоляция, считающаяся одним из главных критериев вида (или даже самым главным), может быстро развиваться под действием отбора как в эксперименте, так и в природных условиях. Возникновение репродуктивной изоляции — ключевой этап видообразования. После того, как между генофондами двух популяций прекратился обмен генами, дальнейшее расхождение (дивергенция) этих популяций становится неизбежной просто в силу статистических факторов. В двух популяциях будут фиксироваться разные мутации, как нейтральные, так и полезные. Рано или поздно это неизбежно приведет к полной генетической несовместимости двух разошедшихся видов. Каждая новая мутация должна быть совместима с остальными генами в генофонде популяции, иначе отбор ее отсеет. Однако мутации, возникающие в одной из популяций, не проверяются отбором на совместимость с генофондом другой популяции. Следовательно, рано или поздно в популяции А закрепится мутация, несовместимая с генофондом популяции Б. В природе на формирование полной генетической несовместимости между двумя изолированными популяциями уходит, как правило, от 2–3 до 10–15 млн лет. Но уже задолго до этого момента разошедшие популяции считаются (и являются) двумя разными видами, потому что реально не скрещиваются в природе, хотя и сохраняют такую возможность.
Промежуточные этапы видообразования
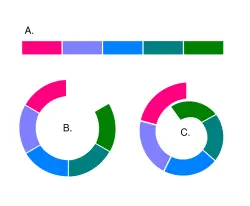
Диаграмма, демонстрирующая варианты скрещивания соседних популяций одного вида. В третьем случае распространение форм может образовать кольцо, в результате две соседние популяции друг с другом не скрещиваются, т. е. формально являются разными видами, хотя и связаны между собой непрерывным рядом взаимно совместимых популяций.
В определении понятия биологического видасущественное значение имеет практически полная репродуктивная изоляцияв природных условиях. Если видообразованиепроисходит постепенно, в результате небольших последовательных изменений, то и сегодня мы должны наблюдать все возможные стадии видообразования, а значит и все возможные стадии репродуктивной изоляции: свободное скрещивание между популяциями, затрудненное скрещивание, снижение плодовитости потомства, бесплодное потомство, и наконец — полную генетическую изоляцию. Если ничего подобного среди современных видов обнаружить не удается, то тем самым опровергаются основы эволюционного учения, то есть это пример фальсифицируемостиэволюционного учения (см.: Speciation stages).
Существует множество примеров того, как разные виды могут скрещиваться в исключительных условиях. Например, у черной вороныи серой вороныразные ареалы, но на пересечении этих ареалов виды могут образовывать гибриды. Еще один пример — различные виды рыбы рода чукучановиз реки Платт, которые живут рядом, но редко скрещиваются.
Неполное видообразование можно наблюдать у различных кольцевых видов: представители вида обитают вокруг гор, водоемов и т. п., при этом соседние популяции скрещиваются между собой и незначительно различаются, а крайние формы различаются значительно и не скрещиваются (см. диаграмму). Пример кольцевого вида — безлёгочная саламандра Ensatina eschscholtzii , обитающая на склонах гор близ тихоокеанского побережья Северной Америки. В зависимости от района обитания вокруг гор саламандры образуют различные формы, постепенно изменяющие свои морфологические и экологические характеристики. Крайние формы Ensatina eschscholtzii eschscholtzii и Ensatina eschscholtzii klauberi , первая из которых имеет красноватую окраску, а вторая чёрно-белую, сосуществуют в узкой зоне гор на юге Калифорнии, однако между собой не скрещиваются.
Еще один пример — серебристая чайка ( Larus argentatus ) и клуша ( Larus fuscus ). Они встречаются в Англии, где их признали разными видами. Но при этом между ними существует цепочка гибридных популяций, идущая вокруг северного полюса. Другие примеры кольцевых видов — зелёная пеночка ( Phylloscopus trochiloides ), оленья мышь ( Реromyscus maniculatus ), наземная улитка ( Partula olympia ), дрозофила Drosophila paulistorum .
У многих гибридов снижена фертильность. У гибридов землероек из рода бурозубок размножение затруднено вследствие разницы хромосомных наборов. Тот же эффект наблюдался в лаборатории при скрещивании Drosophila pseudoobscura из Юты и Калифорнии. Фертильность также снижена у гибридов лягушек из семейства бомбиновых жерлянки краснобрюхой (Bombina bombina) и жерлянки желтобрюхой (Bombina variegata).
Многие другие виды могут давать бесплодное гибридное потомство. В их числе — различные земноводные (например — некоторые виды лягушек из рода Rana ) и млекопитающие (например, мул — гибрид от скрещивания лошади и осла — как правило бесплоден). Гибриды гребенчатого тритона и мраморного тритона бесплодны вследствие наличия непарных хромосом.
При скрещивании ряда других видов достигается оплодотворение, но позднее зародыш погибает. В качестве примера можно привести леопардовую лягушку ( Rana pipiens ) и лесную лягушку ( Rana sylvatica ) из рода настоящих лягушек. Аналогичная ситуация наблюдается у дрозофил и у некоторых растений, таких как хлопчатник обыкновенный ( Gossypium hirsutum ) и хлопчатник барбадосский ( Gossypium barbadense ).
Белый медведь
Конкретным примером крупномасштабных эволюционных изменений является белый медведь( Ursus maritimus ), который, несмотря на родство с бурым медведем( Ursus arctos ), очевидное в силу того факта, что эти виды могут скрещиваться и давать плодовитое потомство, приобрел значительные физиологические различия с бурым медведем. Эти различия позволяют белому медведю комфортно жить в условиях, в которых бурый медведь бы не выжил. В частности, белый медведь способен проплыть десятки километров в ледяной воде, сливается со снегом и не замерзает в Арктике. Всё это возможно благодаря конкретным изменениям: белая окраска способствует маскировке хищника при охоте на тюленей; полые волоски увеличивают плавучесть и сохраняют тепло; слой подкожного жира, толщина которого к зиме доходит до 10 сантиметров, обеспечивает дополнительную теплоизоляцию; удлиненная по сравнению с другими медведями шея позволяет легче держать голову над водой во время плавания; увеличенные лапы с перепонками действуют как весла; небольшие бугорки и полости-присоски на подошвах уменьшают опасность поскользнуться на льду, а плотная шерсть на подошвах защищает лапы от сильного холода и обеспечивает трение; уши меньше, чем у других медведей, и уменьшают потери тепла; веки действуют как солнечные очки; зубы острее, чем у других медведей, и больше подходят для полностью мясного рациона; увеличенный объем желудка позволяет голодному хищнику съесть сразу целого тюленя, кроме того белый медведь способен обходиться без пищи до девяти месяцев за счет переработки мочевины.
Читать дальшеИнтервал:
Закладка:



