Эдвард Стил - Что, если Ламарк прав? Иммуногенетика и эволюция

- Название:Что, если Ламарк прав? Иммуногенетика и эволюция
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Эдвард Стил - Что, если Ламарк прав? Иммуногенетика и эволюция краткое содержание
Что, если Ламарк прав? Иммуногенетика и эволюция - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
В конфигурации зародышевой линии ДНК не транскрибируется в мРНК, по которой могли бы синтезироваться Н- или L- белковые цепочки антитела. В соматической конфигурации перестроенные V(D)J- и С-участки активно транскрибируются, образуется мРНК, которая транслируется в Н- и L- цепи антитела. В неперестроенных V-элементах никогда не происходит соматического мутирования. Мутации появляются только в V(D)J-перестройке.
Перестройки ДНК выполняются набором особых рекомби-национных ферментов. Кодирующие их гены называются генами, активирующими рекомбинацию (recombination activating genes — RAG), и локализованы где-то в других местах генома. RAG-ферменты распознают ДНК-последовательности (реком-бинационные сигнальные последовательности) на соответствующих концах V-, D- и J-элементов, которые и соединяются. Первым шагом в экспрессии тяжелой цепи в созревающей В-клетке костного мозга человека или мыши является DJ-пере-стройка. Вся промежуточная ДНК между D-элементом (их может быть 20 или более) и J-элементом (их 4 или 5) удаляется. Следующий шаг — перемещение случайно выбранного V-эле-мента к перестроенному DJ-элементу, причем снова вся промежуточная ДНК удаляется.
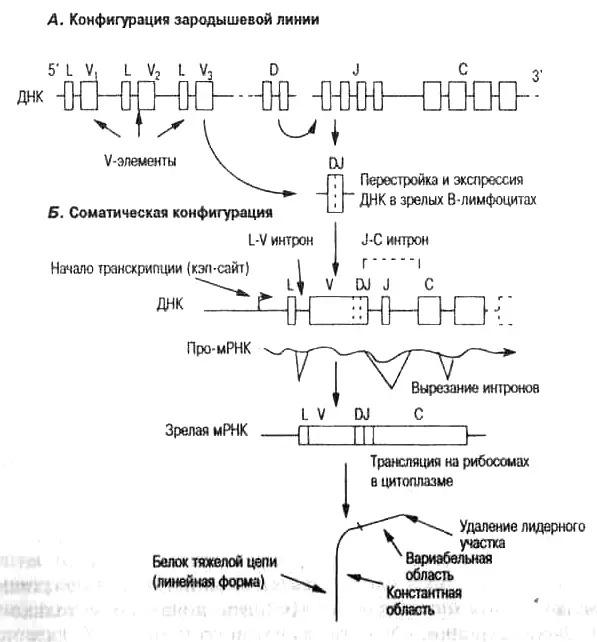
Рис. 4.5. Строение генов, кодирующих V- и С-области тяжелых цепей антитела, в зародышевой линии и после соматической перестройки у человека и мыши.
А. В конфигурации зародышевой линии ряд очень похожих (но все-таки различных) вариабельных V-элементов лежит выше (левее) экзонов, кодирующих константную С-область. Эти V-элементы зародышевой линии (кодирующие примерно 95 аминокислот) называются неперестроенными. Между V- и С- элементами расположены D- и J-элементы (каждый кодирует короткую последовательность аминокислот). Каждый V-элемент имеет лидерную (L) последовательность, которая кодирует короткую последовательность аминокислот, сигнальный пептид. Все гены, кодирующие белки, например, антитела, предназначенные для выведения из клетки (для секреции) или для перехода в другие окруженные мембраной области клетки, имеют такие сигнальные пептиды, которые позволяют им перемещаться в нужные внутриклеточные или внеклеточные области. Интрон между L и V называется L-V-интроном (или лидерным интроном). Его значение для понимания следов интеграции сомы в зародышевую линию подробно описано в гл. 6.
Б. В ходе развития В-клеток в костном мозге в каждом В-лимфоците случайно перестраиваются V-, D- и J-элементы тяжелой цепи, и появляется перестроенная последовательность вариабельной области VDJ, которая теперь называется соматической конфигурацией. Для Н-цепей на первом этапе перестройки происходит объединение DJ, за ним следует объединение с V-элементом, и образование VDJ. После перестройки обнаруживается, что вся ДНК между случайно выбранным V-элементом и использованным J-элементом удаляется из клетки. Таким образом, каждая успешная перестройка ДНК вариабельной области уникальна для каждой зрелой В-клетки. Обратите внимание, что перестроенный участок (VDJ) лежит выше (левее) экзона, кодирующего С-белок. Заметьте также, что в участке между VDJ и С, названном J-C-интроном, лежит неиспользованный элемент (который в этом примере просто удаляется). Про-мРНК, содержащая вариабельный (VDJ) участок, присоединенный к С-участку, образуется в ядре. L-V и J-C интроны (и интроны внутри С -участка) затем вырезаются, что приводит к экспорту зрелой РНК молекулы в цитоплазму, где она транслируется на рибосоме в последовательность аминокислот. Лидерный (сигнальный) пептид отрезается от белка, когда он выделяется из клетки (объяснения терминов см. также в табл. 3.1 и 5.1, на рис. 4.4 и в тексте). Соматическая конфигурация и конфигурация зародышевой линии генов L-цепей такая же, кроме того, что легкие цепи не содержат D-участков.
Продемонстрированные Тонегавой V(D)J-nepecтройки (случайное использование примерно 100 V-генов, 20 D-генов и 4 J-генов и случайное объединение тяжелых и легких цепей для образования гетеродимерного связывающего центра) дают возможность продуцировать большое потенциальное разнообразие белков из сравнительно небольшого количества генетического материала. Кроме того, иногда слияние V, D, и J приводит к добавлению или потере оснований, увеличивая, таким образом, разнообразие.
Разобравшись в этих молекулярных процессах, мы можем теперь задать два очень важных вопроса. Почему В-лимфоцит производит антитела только одной специфичности? Почему соматические мутации (в В-лимфоцитах) возникают преимущественно в перестроенном V(D)J-участке?
Все соматические клетки (клетки тела) имеют двойной набор хромосом, один набор от матери, другой от отца. Такие клетки называются диплоидными. Сперматозоиды и яйцеклетки имеют только по одному набору хромосом (они гаплоидны). Оплодотворение, слияние сперматозоида и яйцеклетки, восстанавливает диплоидное состояние. Кроме половых хромосом (X и Y), все другие хромосомы диплоидного набора разбиваются на пары с очень похожими ДНК-последовательностями. По мере созревания В-лимфоцита начинается перестройка V-, D- и J-элементов. Успешная V(D)J-перестройка в одной хромосоме посылает сигнал, который выключает активность RAG-ферментов и препятствует перестройке в гомологичной хромосоме. Вот, в принципе, ответ на первый вопрос. Дальнейшие детали мы не рассматриваем.
Второй вопрос, почему мутации ограничиваются V(D)J-участком, чрезвычайно важен для обсуждения обратной связи между сомой и зародышевой линией, которая составляет главную тему этой книги. Мы подробно расскажем об этом в следующей главе. Здесь подчеркнем лишь некоторые общие принципы. Если мутации в вариабельной области антитела могут оказаться полезными, то мутации в константной области, скорее всего, вредны, так как они могут нарушить эффекторные функции антитела, запуск лизиса бактериальных клеток или стимулирование фагоцитоза (рис. 3.1). Разделение V- и С-участков позволило эволюции создать механизм, который обеспечивает мутации в V-участке, но сохраняет С-участок неизменным. Вот что происходит в В-лимфоците на определенных стадиях его жизни. Перестроенный V(D)J-ген может подвергаться очень высокому уровню соматических мутаций, и новое антитело оценивается по способности связывать антиген (см. следующую главу). Антитела с самой высокой аффинностью к чужеродному антигену выигрывают в отборочных соревнованиях за связывание антигена, сохраняются и становятся долго живущими клетками памяти. В-лимфоцит проиграет соревнование, если мутация уменьшает или уничтожает аффинность; такие клетки погибают. В-лимфоциты, кодирующие антитела, которые связывают собственные антигены, также уничтожаются, и, следовательно, сохраняется аутотолерантность.
Читать дальшеИнтервал:
Закладка: