Эдвард Стил - Что, если Ламарк прав? Иммуногенетика и эволюция

- Название:Что, если Ламарк прав? Иммуногенетика и эволюция
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:неизвестен
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Эдвард Стил - Что, если Ламарк прав? Иммуногенетика и эволюция краткое содержание
Что, если Ламарк прав? Иммуногенетика и эволюция - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
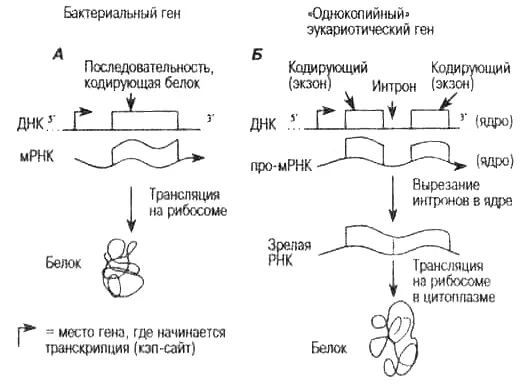
Рис. 4.4. Строение бактериального гена и однокопийного эукариотического гена. На рисунке показаны различия в строении генов, кодирующих белок, у бактерий и у высших клеток. Обратите внимание, что левый конец цепи ДНК принято обозначать как 5'-конец, правый конец — как 3' (произносится как «пять штрих» и «три штрих», соответственно). Слово «однокопий-ный» означает, что в хромосоме находится только одна функциональная копия гена (в отличие от семейства V-генов, рис. 4.5, к которому относится много генов с очень высокой степенью сходства ДНК-последовательностей, расположенных в одной хромосоме). Участки ДНК, которые кодируют белок, изображены в виде прямоугольников. Двухцепочечная ДНК изображается прямой линией, одноцепочечная информационная мРНК — волнистой линией. Изогнутые стрелки указывают на точку начала транскрипции мРНК.
А. У бактерий участок, кодирующий белок, представлен непрерывной рядом триплетных кодонов, каждый из которых определяет одну аминокислоту (см. приложение). мРНК транскрибируется и немедленно транслируется в аминокислотную последовательность (белок) в рибосомах.
Б. У эукариот почти все гены, кодирующие белки, устроены сложнее. У них последовательности, кодирующие белок (экзоны), прерываются некодирующими последовательностями (интронами). На рисунке это обозначено промежутками между кодирующими участками. Редким исключением (гены без интронов) в высших клетках являются гены, кодирующие гистоны (табл. 5.1), а также псевдогены и функциональные ре-трогены, появившиеся в результате обратной транскрипции молекулы мРНК (см. гл. 7). Таким образом, по матрице ДНК создается длинная содержащая интроны молекула про-мРНК. Особая молекулярная внутриядерная органелла (сплайсосома) определяет границы между интронами и экзонами, вырезает интроны из про-мРНК и точно соединяет экзоны, образуя непрерывный кодирующий участок. Затем зрелая, или «процессированная», мРНК экспортируется из ядра в цитоплазму, где она транслируется в последовательность аминокислот в рибосоме.
Молекулярно-генетические исследования Ig-генов начались в середине 1960-х гг. К этому моменту было известно, что и тяжелые (Н), и легкие (L) цепи антитела выполняют двойственную функцию (рис. 3.2). Они состоят из вариабельной аминокислотной последовательности, или V-домена (служит для связывания антигена), и константной, или С -домена (служит, например, для запуска лизиса клетки-мишени или фагоцитоза). Гены антител отличаются от всех остальных генов, локализованных и картированных генетиками в определенных местах хромосом (называемых локусами). Гены, кодирующие V-и С-домены, отделены друг от друга огромным участком последовательности ДНК. Например, V- и С-домены тяжелых цепей мыши находятся в хромосоме 12, около концевого участка длинного плеча хромосомы. Генетическое картирование и последующее изучение последовательности нуклеотидов ясно показали, что V-участок отстоит от С-участка по крайней мере на 300 000 п. н. (300 тыс. п. н.) оснований! Почему?
Даже в середине 1970-х гг., когда стала известна экзоно-ин-тронная структура генов, такое большое расстояние казалось необычным. Однако десятью годами раньше Драйер (Dreyer) и Беннет (Bennet) предположили, что единый участок ДНК, кодирующий Н- или L-цепь, образуется в результате перемещения (транслокации) V-участка к С-участку. То есть, для объяснения удивительного свойства Ig-генов было предсказано существование особого типа редактирования, или сплайсинга ДНК-последовательностей. Далее, эти авторы допускали, что данный процесс происходит при созревании лимфоцита. Драй-ер и Беннет предположили, что часть Ig-локуса (или места локализации в хромосоме последовательности ДНК, кодирующей Ig-гены) содержит в зародышевой линии большое число (сотни) V-генов. В каждой отдельной В-клетке один из этих генов передвигается и становится рядом с одним из нескольких возможных С-генов. Случайное перемещение только одного V-ге-на из большого числа возможных приводит к тому, что клетка производит антитела только одной специфичности. После такой случайной перестройки генов Н- и L-цепей клетка из большого числа возможных комбинаций Н + L белковых цепей создает один полный белок. Затем клетка экспрессирует это антитело на поверхности, и ее судьба решается окончательно: если она окажется «против своих», она будет уничтожена. Если она выжила, она станет потенциальным участником иммунного ответа на чужие антигены.
Это теоретически привлекательное, хотя и достаточно смелое предсказание перестройки ДНК, было доказано лишь десять лет спустя Сусуми Тонегава, который описал организацию и перестройку генов Ig-локуса у мыши[3]. Оказалось, что перестройка ДНК много сложнее, чем предполагалось. Этот процесс изображен на рис. 4.5; он имеет место и у мыши, и у человека. У других позвоночных может быть несколько иная, но в принципе сходная организация кодирующих элементов Ig-локуса.
Несмотря на то, что до точного картирования и секвениро-вания (определения последовательности нуклеотидов) каждого V-элемента генома мыши и человека еще далеко, у нас есть четкая картина строения Ig-генов в клетках зародышевой линии и в соматических клетках. На рис 4.5 приведена схема строения гена тяжелой цепи Ig, но очень похожие схемы можно нарисовать и для генов легких цепей Ig, и для ТкР-генов. Обратите внимание, что около ста V-генов (или V-элементов) расположено левее соединительных J-элементов (от англ. joining — J) и D-элементов (от англ. diversity — разнообразие), а они, в свою очередь, отделены от небольшого числа (примерно 8) генов константной области (C-constant). D- и J-элементы кодируют от трех до пятнадцати аминокислот, тогда как V-элемент — около 100 аминокислот.
О генах антител в неперестроенном виде говорят, что они имеют «конфигурацию зародышевой линии». В таком виде находится ДНК в половых клетках (сперматозоидах и яйцеклетках) и во всех клетках организма, кроме зрелых лимфоцитов (в клетках печени, почек, поджелудочной железы, в других лимфоцитах, например, в фагоцитах). В каждом созревающем лимфоците мыши и человека ДНК подвергается случайной соматической перестройке. Один из V-элементов перемещается и соединяется с одним D- и одним J-элементом. Этот участок располагается перед первым С-геном. Кодирующий С-участок разделен рядом интронов. После этого все еще остается длинная промежуточная последовательность между перестроенным V-геном, который теперь называется V(D)J-ген, и С-участком. Эту структуру называют «соматической конфигурацией». Скобки вокруг D в общем символе гена появляются, потому что гены легких цепей Ig построены только из V- и J-элементов.
Читать дальшеИнтервал:
Закладка: