М. Канунго - Биохимия старения

- Название:Биохимия старения
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:1982
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
М. Канунго - Биохимия старения краткое содержание
Предназначена для биологов, биохимиков, геронтологов, врачей-гериатров.
Биохимия старения - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Если даже рассматривать геном в качестве того первичного участка, где возможны перемены после достижения животным половой зрелости, необходимо иметь в виду, что изменения внеклеточного окружения, связанные с температурой, питанием и т. д., существенно влияют на функцию генома. Поэтому у представителей различных видов и популяций наблюдаются колебания во времени начала старения, его скорости и продолжительности пострепродуктивного периода.
Полученные к настоящему времени данные указывают на то, что старение имеет генетическую основу. Рядом авторов разработаны теории, в которых делаются попытки объяснить старение на уровне генома. Ниже мы обсудим теории только этого типа.
Росс и Скотт [104] первыми сообщили, что крысы, подвергнутые тотальному облучению, слишком слабому, чтобы вызвать какие-либо острые изменения, погибают раньше, чем необлученные контрольные животные. Затем последовали сообщения о том, что у облученных грызунов [26, 44, 108] и людей [128] симптомы старения, и смертность были такими же, как у интактных особей; было отмечено, однако, что частота опухолей у первых была выше. Поэтому предположили, что облучение вызывает ускорение процесса старения. Основываясь на этих данных, Сцилард [122, 123] предложил для объяснения старения "теорию соматических мутаций", согласна которой мутации, возникающие беспорядочно и самопроизвольно, разрушают гены и хромосомы постмитотических клеток в течение жизни организма, постоянно повышая мутационный груз. При увеличении числа мутаций и потере функциональных генов наблюдается снижение синтеза функциональных белков. Смерть клетки наступает тогда, когда мутационный груз превышает критический уровень. В результате число постмитотических клеток уменьшается, а общая функциональная активность организма снижается.
Эта теория была проверена в опытах с мышами Стивенсоном совместно с Кёртисом [118] и Кёртисом [20, 21]. Мышей облучали дозой 400 рад или им вводили химические мутагены типа азотистых аналогов иприта (0,125 мг) и затем определяли их выживание. В регенерирующей печени молодых и старых мышей исследовали также частоту хромосомных аберраций типа нерасхождения сестринских хроматид в митозе и хромосомных разрывов. У мышей из линий с различной продолжительностью жизни вызывали частичный некроз печени введением четыреххлористого углерода и подсчитывали число хромосомных аберраций после фиксации в метафазе под действием колхицина.
Было показано, что а) сокращение жизни после рентгеновского облучения зависит от дозы, б) в регенерирующих клетках печени облученных мышей повышена частота хромосомных аберраций (рис. 9.1), в) азотистые аналоги иприта не оказывают влияния ни на продолжительность жизни, ни на частоту хромосомных аберраций, г) в клетках печени короткоживущих мышей линии A/HEJ (395 дней) накапливается больше хромосомных аберраций, чем у долгоживущих мышей линии C57BL/6J (600 дней; рис. 9.2) [19]. Кёртис [20] далее предположил, что проникающая радиация повреждает постмитотические клетки больше, чем премитотические. В первых мутационные эффекты аккумулируются и клетка не может устранить их, так как она не делится. Во втором случае поврежденная клетка элиминируется и замещается неповрежденной клеткой. Клетки зародышевого пути более устойчивы к повреждению хромосом, благодаря чему возможно сохранение вида.
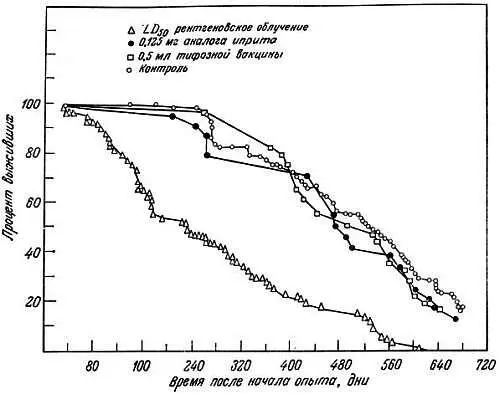
Рис. 9.1. Кривые выживания мышей после различных воздействий, начиная с 2-месячного возраста [20]. Кривые начинаются через 30 дней после воздействия — срока, достаточного для того, чтобы не учитывать внезапную гибель. Они показывают, что однократные массивные, но не смертельные дозы ядовитых химических веществ не уменьшают продолжительности жизни, тогда как единичные массивные, но не смертельные дозы рентгеновского облучения дают заметный эффект
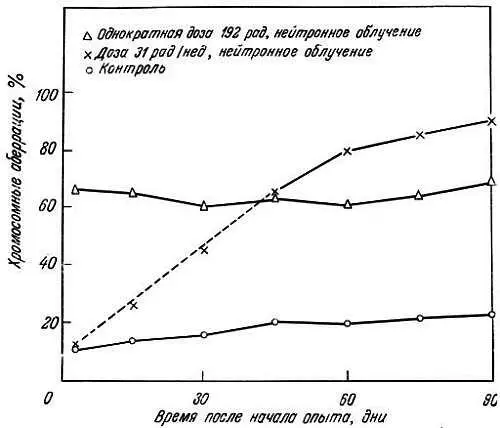
Рис. 9.2. Сравнение эффекта нейтронного облучения на число хромосомных аберраций в клетках печени мышей и их возрастных изменений [22]. При однократном облучении дозой 192 рад аберрации наблюдаются в среднем) в 56 % клеток (треугольники)
Кларк и Рубин [17] изучили действие рентгеновского излучения на продолжительность жизни бабочки Habrobracon , у которой самки диплоидны, а самцы либо диплоидны, либо гаплоидны. Интактные самцы, как диплоидные, так и гаплоидные, имеют одинаковую продолжительность жизни. Однако гаплоидные особи более чувствительны к проникающей радиации, чем диплоидные. Это показывает, что соматическая мутация не может быть причиной старения; если бы это было так, то диплоидные особи имели бы большую продолжительность жизни, а диплоидные самцы и самки имели бы одинаковую продолжительность жизни. Диплоидные самцы более устойчивы к радиации. Это говорит о том, что повреждение, вызываемое облучением, восстанавливается более эффективно, если число хромосом больше, и что процесс восстановления не зависит от пола. Итак, снижение продолжительности жизни, вызванное радиацией, отличается от естественного старения. Здесь уместно процитировать Хендлера [42]: "Представление о том, что облучение приводит к преждевременному старению, верно наполовину и в широком смысле отражает нечеткость исходного параметра — продолжительности жизни".
Аналогичные наблюдения были проведены Томпсоном и Холлидеем [125] на уровне клеток in vitro. Когда фибробласты легких эмбриона человека (штамм MRC-5) обрабатывают колхицином в течение 3–6 ч, выживающая популяция содержит около 60 % тетраплоидных клеток, которые продолжают делиться. Продолжительность их жизни не отличается от продолжительности жизни диплоидных клеток. Сопоставима и скорость их роста. Если бы причиной смерти клеток было накопление мутаций или генетические дефекты, то тетраплоидные клетки должны были бы обладать большей устойчивостью и имели бы большую продолжительность жизни. Но это не так. Хён и др. [46] также наблюдали, что диплоидные фибробласты кожи человека имеют ту же продолжительность жизни, что и тетраплоидные. Если мутации и вносят свой вклад в процессе старения клеток в культуре, они не проявляют себя, в период активного роста (фаза II) и потому должны быть рецессивными. Возможно, что происходит постепенное увеличение генетического груза рецессивных дефектов, что может в итоге привести к инактивации одного или нескольких необходимых генов обеих гомологичных хромосом. Если это так, то тетраплоидные клетки могли бы противостоять большим повреждениям и имели бы большую жизнеспособность. Если мутации вредны, они вызывают гибель клеток и не аккумулируются. Диплоидные и тетраплоидные клетки, имели бы сходную продолжительность жизни только в том случае, если бы вредные мутации возникали в конце жизни.
Читать дальшеИнтервал:
Закладка:









