М. Канунго - Биохимия старения

- Название:Биохимия старения
- Автор:
- Жанр:
- Издательство:неизвестно
- Год:1982
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
М. Канунго - Биохимия старения краткое содержание
Предназначена для биологов, биохимиков, геронтологов, врачей-гериатров.
Биохимия старения - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Во-вторых, хотя на ранних стадиях развития репликация ДНК, а затем деление клеток происходят во всех клетках, после некоторого периода увеличения числа клеток и развития организма на определенных стадиях дифференцировки в клетках некоторых типов синтез ДНК и деление клеток прекращаются. В качестве примера можно привести нейроны, а также клетки скелетной и сердечной мышцы позвоночных, которые перестают делиться вскоре после рождения, т. е. становятся постмитотическими. Некоторые из них по окончании периода развития стареют и умирают, но большая часть продолжает функционировать в течение всей жизни. Так, в клетках костного мозга, эпителия и т. п. синтез ДНК и деление продолжаются на протяжении всей жизни, т. е. эти клетки остаются премитотическими.
Каково же значение этих двух функциональных изменений в ДНК для организма и для процесса старения? Все многоклеточные организмы начинают стареть после достижения половой зрелости. Являются ли причиной старения дифференцировка и (или) постмитотическая природа клеток? Будет ли предотвращено старение, если остановить одно или оба изменения ДНК? Являются ли эти функциональные изменения ДНК необратимыми? Известно, что ДНК в клетках не находится в изолированном состоянии. Она связана в комплекс с белками двух типов: гистонами и негистоновыми хромосомными белками (НГБ), которые вместе с ДНК образуют надмолекулярный комплекс, называемый хроматином и представляющий собой генетический аппарат эукариотов. Три компонента присутствуют в комплексе приблизительно в равных пропорциях. Здесь же обнаружена и РНК, однако полагают, что она является продуктом транскрипции ДНК, а не структурным компонентом. Функция ДНК известна, роль же белков в функционировании хроматина определена недостаточно. Изменяются ли они в течение жизни? Для того чтобы выяснить, вносят ли вклад в процесс старения изменения в одном или нескольких компонентах хроматина, необходимо установить его химический состав и структуру. Структура и функции хроматина описаны в нескольких обзорах [12, 57, 74, 112, 116, 199,354].
Гистоны — белки с малой молекулярной массой — обнаружены в хроматине всех эукариотов. Их впервые открыли в 1943 г. Стедман и Стедман [330]. Эти белки имеют основной характер и положительно заряжены при физиологических значениях рН, поскольку они богаты лизиновыми и аргининовыми остатками. Они не содержат триптофана и присутствуют в клетках в отношении 1:1 с ДНК. Имеется пять основных типов гистонов: Н1, H2A, H2B, Н3 и Н4, которые различаются по величине соотношения лизина и аргинина. Их легко разделить с помощью электрофореза в полиакриламидном геле (рис. 2.1). Некоторые характеристики гистонов из тимуса теленка приведены в табл. 2.1.
Таблица 2.1. Параметры гистонов из тимуса теленка
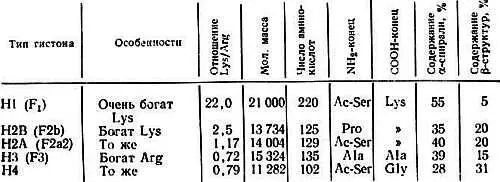
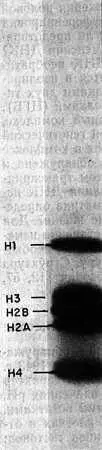
Рис. 2.1. Электрофореграмма гистонов в полиакриламидном геле
Важное свойство всех гистонов состоит в том, что их положительно заряженные лизиновые и аргининовые остатки образуют кластеры в особых областях полипептидной цепи. Этим и объясняется наличие во вторичной структуре гистонов вытянутых β-структур. Очевидно, эти положительно заряженные β-структуры связываются с отрицательно заряженными фосфатными группами ДНК сильнее, чем с другими группами. Нейтрализация положительных зарядов в гистонах должна приводить к их отделению от ДНК. Из табл. 2.1 видно, что гистоны H2A, H2B, Н3 и Н4, находящиеся внутри нуклеосом, имеют больше вытянутых β-структур, чем гистон Н1, расположенный между нуклеосомами.
Прокариоты не имеют гистонов. Появление гистонов совпало с возникновением ясно выраженных ядер, хромосом и процесса дифференцировки. Гистоны подавляют синтез РНК [167] и ДНК [142] in vitro. При полном или частичном удалении гистонов из хроматина сильно увеличивается его матричная активность. Однако маловероятно, чтобы гистоны контролировали или регулировали транскрипцию генов, так как: а) имеется только пять основных видов гистонов, тогда как геном эукариотов содержит несколько тысяч генов; б) количество гистонов обычно постоянно в клетках всех типов и во всех периодах жизни; в) количество гистонов одинаково в метаболически активных и метаболически неактивных клетках. Следовательно, можно предположить, что гистоны включены в структуру и организацию хроматина и действуют как общие репрессоры его активности.
Гистон Н1
Гистон Н1 очень богат лизином — около 25 % входящих в его состав аминокислотных остатков составляет лизин. Он отделяется от ДНК гораздо легче других гистонов. Этому гистону свойствен полиморфизм, т. е. в одной ткани может быть несколько видов гистона Н1 с различными последовательностями аминокислот. В тимусе и печени крыс обнаружены пять изогистонов Н1. Относительное содержание изогистонов Н1 в разных тканях одного и того же организма различно [58, 114, 189, 190] и изменяется в течение клеточного цикла [160]. Показано, что различные подфракции гистона Н1 морского ежа синтезируются на разных стадиях развития яйца. В некоторых яйцах одна фракция гистона Н1 заменяется на другую во время перехода от бластулы к гаструле; в других это изменение происходит во время вылупления. Разные фракции гистона Н1 из тимуса кролика дают различные спектры кругового дихроизма с ДНК фага Т7 [370]. Отсюда следует, что различные подфракции гистона Н1 имеют различные функции [8, 307].
При изучении последовательности аминокислот подфракций гистона Н1 было показано, что в отличие от остальных четырех гистонов он имеет основной концевой COOH-участок. Концевая NH 2-область (1-40) также имеет основной характер (24–39). В концевой NH 2-области изогистонов Н1 найдено большое число аминокислотных замен. Эти замены, по-видимому, определяют функциональные различия изогистонов Н1 во взаимодействии с НГБ и эффекторами, а также в связывании с ДНК. Концевая NH 2-область представляет собой неупорядоченную спираль. Центральный участок [(39±4)-(116±4)] кроме большого числа аминокислот кислотного характера и двух ароматических аминокислот содержит неполярные аминокислоты. Этот участок способен к образованию вторичной глобулярной структуры. Он в основном инвариантен и весьма консервативен, т. е. все гистоны Н1 различных организмов имеют в этой области практически одну и ту же последовательность аминокислот. По-видимому, она играет существенную роль в структуре хроматина.
Концевая COOH-область является сильно основной из-за наличия большого числа лизиновых остатков и весьма консервативна внутри одного вида. Поэтому она может играть общую роль во всех гистонах Н1. Она также представляет собой неупорядоченную спираль. Эта область в основном ответственна за связывание с ДНК. Предполагают, что основные области гистона Н1 связываются с ДНК, а неполярная и глобулярная центральная область взаимодействует с другими молекулами [77, 89, 154]. Стафилококковая дезоксирибонуклеаза специфически расщепляет хроматин между нуклеосомами, в результате чего образуются фрагменты ДНК, связывающие две соседние нуклеосомы. Показано, что гистон Н1 соединяется приблизительно с 30–60 парами оснований этих фрагментов ДНК, т. е., по-видимому, гистон Н1 не участвует в образовании структуры нуклеосомы, а располагается в областях между нуклеосомами. Положительный заряд гистона Н1 выше, чем у других гистонов. Он первым вытесняется из хроматина кислотой или щелочью и в большей степени подвержен разрушению протеазами, когда еще находится в связанном состоянии в комплексе хроматина [26, 263]. Если гистон Н1 добавить к хроматину с недостаточным содержанием этого гистона, то хроматин сжимается [46, 47]. Если же гистон Н1 смешать с двухцепочечной ДНК, то образуются структуры, имеющие форму бублика (тора) [166]; другие гистоны в подобных условиях участвуют в образовании глобул, похожих на нуклеосомы. Таким образом, гистон Н1, вероятно, участвует в образовании структур хроматина высшего порядка, а именно способствует закручиванию нитей нуклеосом в сверхспиральный виток с диаметром ~20 нм [47]. Аналогичные структуры образует с ДНК гистон Н5. Вероятно, различные подфракции гистона Н1 могут быть связаны с различными межнуклеосомными (линкерными) областями хроматина и участвуют в образовании разных сверхспиралей. В интерфазном хроматине ДНК свернута в несколько тысяч раз, благодаря чему она умещается по длине метафазной хромосомы. Определенную роль в этой конденсации ДНК может играть гистон Н1.
Читать дальшеИнтервал:
Закладка:









